

Вторичные, ассоциационные, или интегративные, поля; 6—лобная область; 6—височно-теменная область (ill—IV—у насекомоядных, V—VI—у полуобезьян).
базальный ганглий с дорзальной и латеральной поверхности. Эта пластинка входит у вышестоящих животных (до млекопитающих включительно) в состав сосудистых сплетений боковых желудочков. Лишь передний участок пластинки на месте эмбрионального переднего отверстия несколько утолщён и называется концевой пластинкой—lamina terminalis. Примитивный плащ первоначально представлен только концевой пластинкой, в которой располагается довольно значительная передняя спайка—commissura nasalis,— состоящая из волокон, соединяющих оба полосатых тела, а впоследствии и обе грушевидные доли. Отсюда следует, что примитивный плащ находится в связи с органом обоняния.
У рептилий (рис. 159 и 160—D, 9) в плаще впервые появляется поверхностно расположенное серое мозговое вещество определённой структуры в виде коры большого мозга. Однако это серое вещество не имеет ещё характера, присущего плащу млекопитающих, и является только высшим обонятельным центром. В силу сказанного считают, что у рептилий плащ первичный—archipallium s. primordium hippocampi—в отличие от вторичного плаща млекопитающих—neopallium.
С разрастанием вторичного плаща (рис. 165—/, //, а, Ь), в котором сосредоточиваются все центры, кроме обонятельных, вентрально лежащий первичный плащ отодвигается в медиальные стенки полушарий и при этом глубоко вдавливается в боковые желудочки, увлекая за собой примитивную
774
НЕРВНАЯ СИСТЕМА
эпителиальную пластинку, участвующую в образовании сосудистого сплетения бокового желудочка. Линия вдавления называется медиальной пограничной щелью—fissura hippocampi (рис. 180—11),—а сама складка первичного плаща, втиснутая в полость желудочка, называется аммоновым рогом— cornu Ammonis s. hippocampus (рис. 181—14),—-который сохраняет свою функцию обонятельного центра. Базальная часть первичного плаща остаётся на базальной поверхности полушарий в виде грушевидных долей—lobi piriformes; им также присуща обонятельная функция.
Продолжающееся разрастание полушарий вторичного плаща обусловливает изменение в положении аммонова рога: так как полушария ложатся на аммонов рог сзади, то, увеличиваясь в размере, они оттесняют каудальные концы рога вентрально и вперёд, в полость грушевидных долей.
В связи с образованием первичного плаща от передней спайки уже у двудышащих рыб и амфибий обособляется спайка плаща, или спайка аммонова рога—commissura pallii anterior»s. commissura hippocampi. В ней проходят коммиссуральные волокна, соединяющие друг с другом половины первичного плаща. Эти спайки у рептилий, птиц ж млекопитающих находятся впереди межжеяудочковых отверстий. По мере разрастания вторичного плаща от передней спайки его обособляются новые коммиссуральные волокна, образующие мозолистое тело, или спайку обоих полушарий— corpus callosum (рис. 181—<5).
Другая часть волокон передней спайки плаща, соединяющая аммоновы рога, также сильно разрастаясь, образует свод над третьим желудочком-fornix (рис. 183—4)—и ножки свода—crura fornicis,—переходящие на акмо-новы рога. Назально к своду присоединяются волокна, направляющиеся в сосцевидное тело. Из этих волокон получаются столбы свода—columnae fornicis.
Между сводом и мозолистым телом сохраняется в виде прозрачной перегородки—septum pellucidum (рис. 183—3)—вторично изменённая часть первичной концевой пластинки—lamina terminalis,—в которой лежит первичное серое ядро.
Борозды плаща
У большинства млекопитающих поверхность вторичного плаща покрыта бороздами и извилинами, вследствие чего такие животные называются гирэнцефалам и—gyrencephali—в отличие от лиссэнцефа-л о в—lissencephali,—у которых борозды и извилины на плаще отсутствуют (рис. 160—G и F).
Благодаря извилинам увеличивается поверхность коры мозга.
Очень распространено убеждение, что мозг человека имеет самое большое количество извилин, однако это не совсем верно: на мозге слонов и китов извилин ещё больше. Извилин вообще больше у крупных животных, в то время как все мелкие животные—лис-сэнцефалы; например, у уистити (самой маленькой обезьяны) извилины и борозды отсутствуют, хотя относительный вес мозга у неё значительно больше, чем у гориллы (1:20— у первой и 1:200—у второй).
В процессе эмбрионального развития борозды появляются в известной последовательности. По ходу борозд и самих извилин мозг млекопитающих (гирэнцефалов) подразделяют на три типа, поэтому абсолютная гомологиза-ция извилин между различными отрядами животных представляет очень большие затруднения, а в некоторых случаях, невидимому, совсем невозможна.
На поверхности головного мозга, наиболее просто устроенной у собаки (рис. 180—/), различают пограничные и главные борозды.
РАЗВИТИЕ И СТРОЕНИЕ ГОЛОВНОГО МОЗГА 775
Назальная пограничная, или обонятельная, борозда—sulcus rhinalis (1)—проходит по латеро-вентральной поверхности головного мозга и отделяет лежащий базально-обонятельный мозг от neopallium. Борозда тянется от обонятельной луковицы каудально,. вдоль латерального края латеральной обонятельной извилины—gyrus olfactorius lateralis—и латеральной (сильвиевой) бороздой (2) делится на назальную и каудальную части. Каудалъная часть (I1) переходит на затылочную долю мозга, образуя затылочно-височную* борозду—sulcus occipitotemporalis (22),—кбторая на медиальной поверхности полушарий часто переходит или только соединяется с sulcus splenialis (см. ниже).
Медиальная пограничная щель—fissura hippocampi (И)—образует на медиальной поверхности полушарий медиальную границу грушевидной доли.
К главным бороздам относятся: 1) латеральная, или сильвиева, борозда— sulcus lateralis Sylvii (2)\ она начинается от базальной пограничной борозды, как раз. в том месте, где на базальной поверхности помещается поперечная ямка—fossa transversa. В сильвиевой борозде проходит средняя мозговая артерия.
Над сильвиевой бороздой располагаются три параллельные дугообразные борозды.
1) Первая дугообразная, или эктосильвиева, борозда—sulcus ectosylvius (4)—огиба ет сильвиеву борозду спереди, дорзально и сзади, вследствие чего её и подразделяют на три части: назальную, среднюю и каудальную.
Отдельные части первой дугообразной борозды (или вся она в целом) у животных вследствие углубления сильвиевой борозды погружаются в последнюю и, таким образом, исчезают с поверхности мозга.
2) Вторая дугообразная, или надсильвиева, борозда—sulcus suprasylvius (5)— идёт параллельно и снаружи от первой. Она также делится на назальную, среднюю и каудальную части, причём каудалъная часть нередко обособляется.
Третья дугообразная, или эктомаргинальная, борозда—sulcus ectomarginalis (б)—идёт параллельно двум первым и дорзальному краю полушарий. Она подразделяется на задний участок—л атеральн ую борозд у—sulcus lateralis (в)—и передний участок—в енечную борозду —sulcus coronalis (б1),—которые могут и не соединяться друг с другом. От заднего конца венечной борозды отходит медиально петлистая борозда—sulcus ansatus (62).
В передней части плаща впереди от дугообразных борозд лежит пресильвиева борозда—sulcus praesylvius (7).
5 и 6) В затылочно-теменной части мозга проходят ещё две борозды: одна медиально от латеральной борозды—sulcus entolateralis [8), а Другая латерально—sulcus ectolate-ralis (9).
7) Передняя часть полушарий от задней отделяется крестовидной бороздой—sulcus cruciatus (10). Эта борозда переходит с латеральной поверхности полушарий на медиальную и типична только для отряда мясоядных—carnivora.
8 и 9) На медиальной поверхности полушарий вдоль мозолистой спайки проходит борозда мозолистого тела—sulcus corporis callosi (12). Этой борозде параллельна поясная борозда—sulcus cinguli s. sulcus callosomarginalis (IS),—которая у домашних животных обычно состоит из двух частей: передней—борозды колена—sulcus genualis (IS1) —и задней—борозды валика—sulcus splenialis (IS).
Деление плаща на доли
Для удобства изучения плащ принято делить на ряд долей.
Лобная доля—lobus frontalis—у собаки помещается впереди sulcus coronalis et ansatus и sulcus Sylvii,—а у свиньи, рогатого скота и лошади—приблизительно впереди плоскости, проходящей через sulcus genualis. Затылочная доля— lobus occipitalis—находится каудально от сегментальной плоскости, проведённой через валик мозолистого тела. Теменная доля—lobus parietalis—лежит между лобной и затылочной долями. Височная доля—lobus temporalis—ориентировочно занимает место между sulcus. Sylvius, sulcus* rhinalis и sulcus suprasylvius. Обонятельная доля—lobus olfactorius и lobus falciformis— образуют обонятельный мозг (ср. рис. 165).
Строение коры большого мозга
Вторичный плащ построен из серого и белого мозгового вещества. Первое располагается поверхностно и образует кору большого мозга, второе находится в глубине.
Кора головного мозга в различных её участках гистологически устроена не одинаково, что впервые было установлено В. А. Бецом (1874) и им же объяснено: «Эта структурная диференциация мозговой коры есть выражение локализированности функций в коре» (1881). Различие касается как
776
НЕРВНАЯ СИСТЕМА
расположения клеточных элементов—ц итоархитектоники, так и хода волокон в сером мозговом веществе—м иэлоархитектоники. Цито- и миэлоархитектоника коры большого мозга более или менее подробно изучены лишь у человека и довольно слабо у животных.
Клеточные элементы коры располагаются параллельно поверхности мозга в шесть слоев (пластов). Пласты эти, считая снаружи внутрь, следующие: /—молекулярный, //—наружный зернистый.. ///—слой малых пирамидных клеток, IV—внутренний зернистый, V—слой больших пирамидных клеток и VI—слой полиморфных клеток (рис. 166). В филогенетическом

Рис. 166. Схема распределения клеток (цитоархитектоника) и волокон (миэлоархитектоника) коры большого мозга.
Слева: I—молекулярный слой; /7—наружный зернистый слой; III—слой малых пирамидных клеток; IV—внутренний зернистый слой; V"—слой больших пирамидных клеток; VI—слой полиморфных клеток; VII—белое мозговое вещество. Справа: /—тангенциальный слой; II—дисфиброзный слой; ///—lamina suprastriata; IV—наружная * байяжеровская полоска; V—lamina interstriata и внутренняя байяжеров-ская полоска; V/—lamina infrastriata.
ряду животных не все шесть слоев бывают резко разграничены, но наличие их можно доказать у всех млекопитающих.
Клетки, образующие кору, делятся на три типа: Р а м о н-К аха-левские, пирамидные и полиморфные. Из них особого внимания заслуживают пирамидные клетки—клетки Беца: чем выше по своей организации или старше животное, тем больше у них отростков, следовательно, и связи этих клеток с другими обширнее (рис. 167).
Различным слоям клеток приписывают различные функции: внутреннему зернистому слою (IV), являющемуся первичным, принадлежит рецеп-торная функция, клеткам V и VI слоев, большим пирамидным и веретенообразным клеткам—эффекторная. Позже появляющиеся клетки 7/ и III слоев выполняют ассоциационные функции высшего порядка, т. е. психические.
На основе местных различий в цитоархитектонике кору млекопитающих животных разделяют на главные области—поля—area,—причём каж-
РАЗВИТИЕ И СТРОЕНИЕ ГОЛОВНОГО МОЗГА
777
дое поле, по крайней мере у высших млекопитающих и особенно у человека, на основе различной миэлоархитектоники может быть подразделено ещё на ряд миэлоархитектонических участков.
Филогенетически все поля во вторичном плаще диференцируются из примитивных четырёх областей низших млекопитающих (сумчатых и насекомоядных); эти области различны по своей цитоархитектонике, и, судя по наличию в них определённых проекционных волокон (рис. 165), им также соответствуют специальные функциональные центры. Так, в лобной области
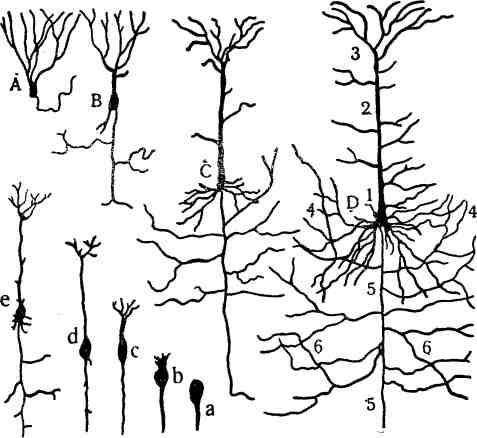
Рис. 167. Схема фило- и онтогенеза пирамидной клетки:
Л—у лягушки, В—у ящерицы, С—у млекопитающего,
D—у человека. f
/—тело клетки; 2—дендрит; 3—его отростки; 4—дендриты боковых поверхностей; 5—нейрит; б—его коллатерали (боковые ветви); fl,ft,c,d, e—стадии эмбрионального развития пирамидной клетки.
лежит примитивный моторный корковый центр, в затылочной—зрительный центр, между ними—кожно-чувствующий, в височной области—слуховой центр. Физиологические и архитектонические центры не всегда соответствуют расположению извилин.
Соотношение мякотных волокон в коре мозга до известной степени определяется принятым делением их на группы соответственно шести слоям клеток (рис. 168, справа).
По развитию миэлиновой оболочки (миэлогенетически) все нервные волокна относят или к проекционной системе, или ассоциационной. Первые развиваются уже до рождения, вторые—только после рождения. Миэлогенетически кора также может быть разделена по двум признакам—процентному соотношению проекционных и ассоциационных волокон и их онтогенезу—на области: первичную, конечную и промежуточную, т. е. волокна, одевающиеся миэлиновой оболочкой до рождения, после рождения или в последний период эмбрионального состояния.
В настоящее время, на основании изучения цито- и миэлоархитектоники коры мозга, у человека выделяют более 250 полей.
Среди волокон, образующих белое вещество мозга, различают ассоциа-ционные, коммиссуральные и проекционные.
778
НЕРВНАЯ СИСТЕМА
Ассоциационные волокна соединяют друг с другом многочисленные участки коры внутри одного полушария. Они делятся на короткие и длинные; первые связывают соседние извилины, вторые—более удалённые друг от друга участки и доли. Длинные волокна особенно сильно развиты у человека, в то время как у животных они или незначительны, или совсем отсутствуют.
Коммисеуралыше волокна соединяют участки, лежащие в разных полушариях. Они образуют переднюю и заднюю коммиссуры (спайки)—спайку аммонова рога и мозолистое тело.
Проекционные волокна анатомически делятся на короткие и длинные, а функционально—на кортико-фугальные и кортико-петальные. Короткие проекционные волокна соединяют кору с ядрами зрительных бугров и четверохолмия. Длинные пути связывают кору с ядрами моста или со спинньщ. мозгом. Все эти пути, как короткие, так и длинные, проходят между хво статым телом и ядрами зрительных бугров, с одной стороны, и чечевичным; ядром, с другой, образуя внутреннюю капсулу—capsula interna (рис. 184—9). На среднем мозге эти волокна формируют ножки большого мозга—pedun-culi cerebri.
Проекционные пути, так же как и ассоциационные, наиболее сильно-развиты у человека; у животных некоторые из них даже совсем отсутствуют.
Таким образом, полушария большого мозга в целом и кора полушарий в частности устроены наиболее сложно и притом не одинаково в ряду млекопитающих, достигая максимума своего развития у высших приматов и особенно у человека. По И. П. Павлову, на большие полушария беспрерывно падают бесчисленные раздражения как из внешнего мира, так и из внутренней среды самого организма. Так как кора мозга высших животных является носительницей замыкательной функции (т. е. функции приобретения, образования новых связей между организмом и средой, развития нового жизненного опыта, функции онтогенетической адаптации, приспособляющей организм к условиям среды), высший отдел центральной нервной системы держит в своём ведении все явления, происходящие в теле.