

ответы на экзамен 2019 г
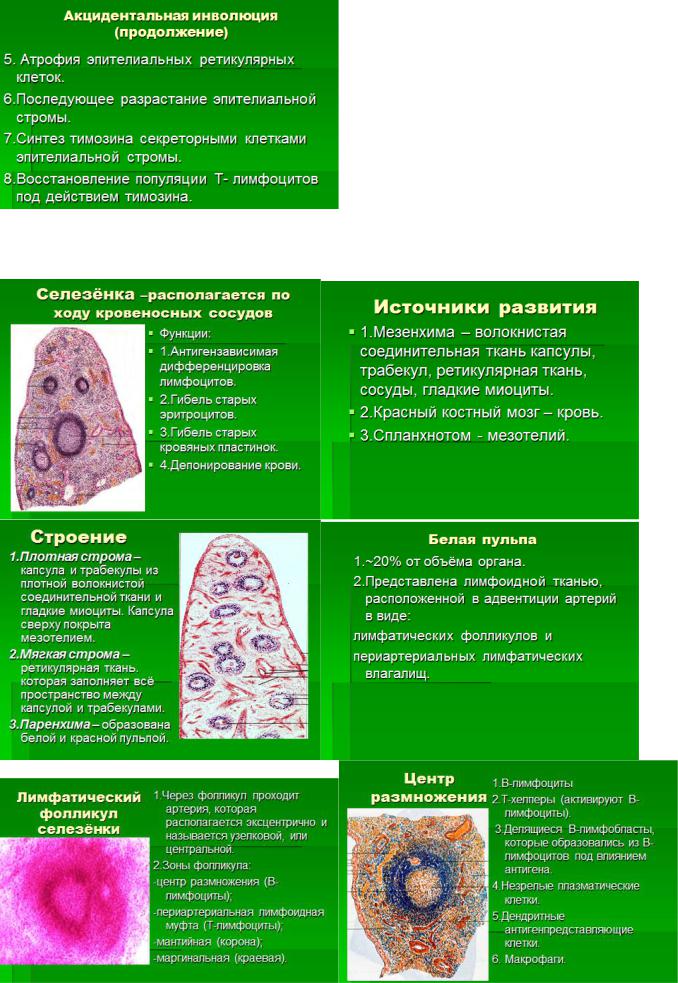
4. Селезенка. Источники развития. Строение красной и белой пульпы. Морфо-функциональная характеристика зон лимфоидных узелков. Стромальные элементы микроокружения. Особенности кровоснабжения в органе. Основные функции селезенки.
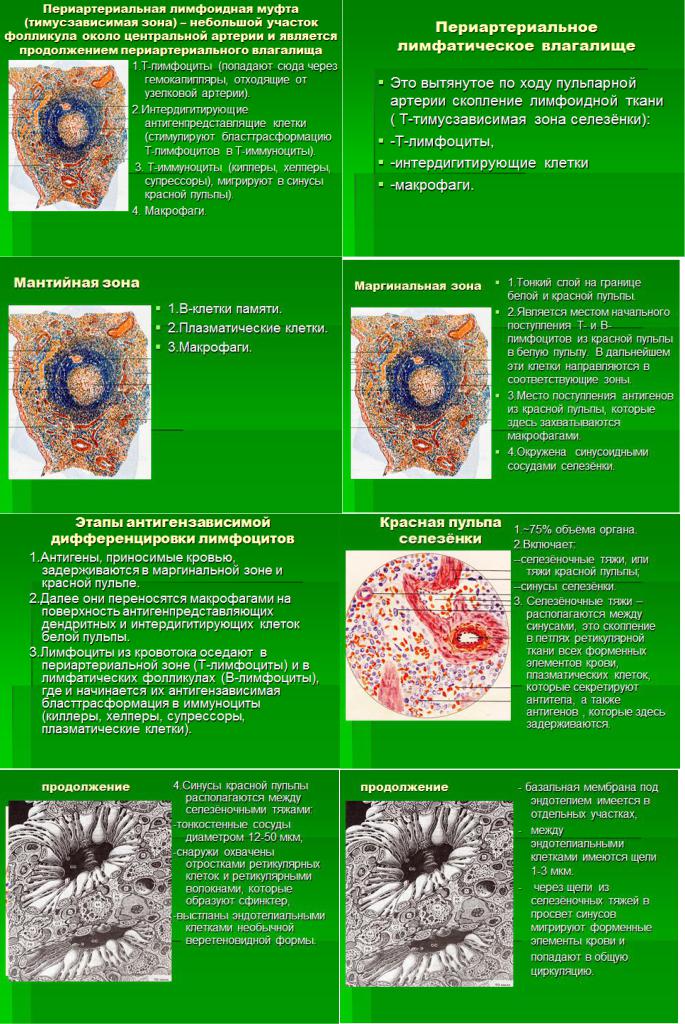

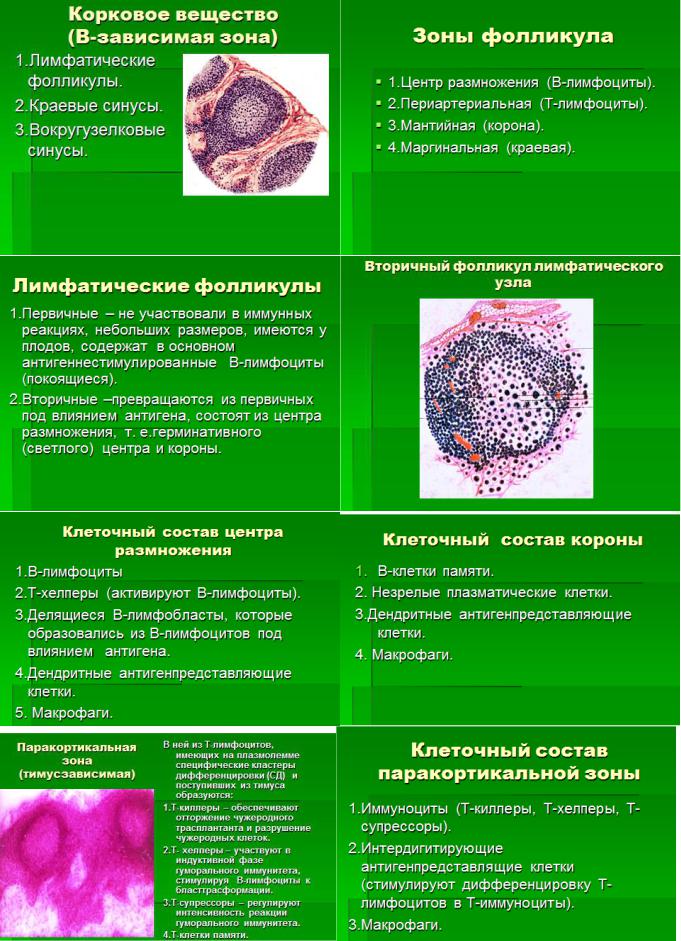
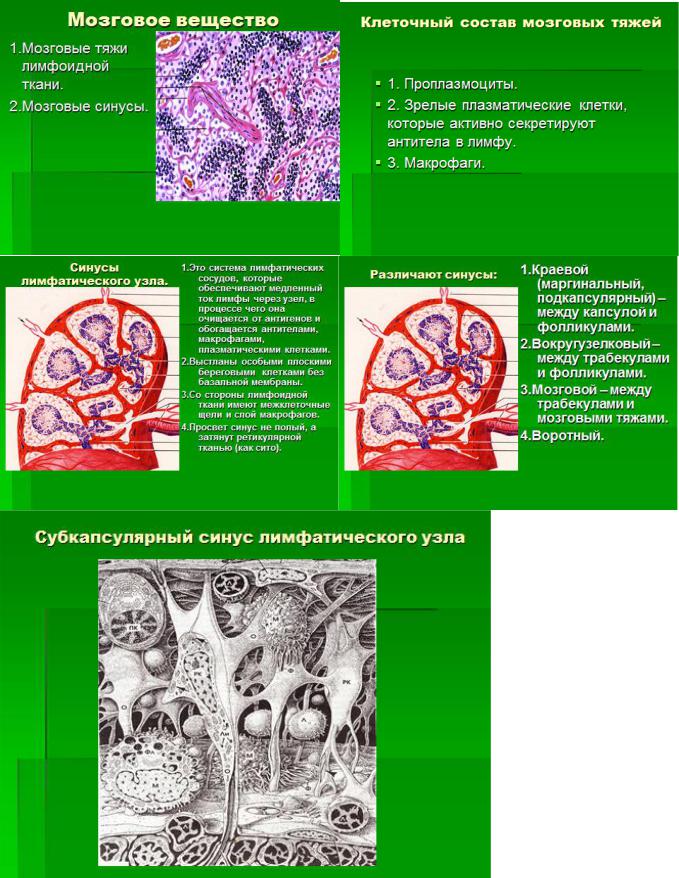
5. Лимфатические узлы. Источники развития. Строение и морфо-функциональные зоны. Стромальные элементы микроокружения. Система синусов. Основные функции лимфатических узлов. В-лимфоциты: субпопуляции и их функции.
6. Понятие о иммунной системе слизистых оболочек и ассоциированной со слизистой лимфоидной ткани (MALT). Небные миндалины. Источники развития. Строение и морфо-функциональные зоны. Стромальные элементы микроокружения. Основные функции миндалин. В-лимфоциты: субпопуляции и их функции. * Морфофункциональные особенности миндалин в раннем детском возрасте
Лимфоидная т ань, ассоциированная со с изистыми обо оч ами |
|
Данный вид имфоидной т ани распо агается под с изистой обо оч ой. Сюда относятся аппенди с, |
имфоидное |
о ьцо, имфатичес ие фо и у ы ишечни а, а та же аденоиды. С оп ения имфоидной т ани в |
ишечни е – |
пейеровы б яш и. |
Эта имфоидная |
т ань яв яется барьером на пути прони новения ми робов через с изистые |
обо оч и. Фун ции |
имфоидных с оп |
ений в ишечни е и минда инах: |
1) распознавате ьная – общая п ощадь поверхности минда ин у детей очень бо ьшая (почти 200 см2). На этой п ощади происходит постоянное взаимодействие антигенов и ето иммунной системы. Именно отсюда информация о чужеродном агенте с едует в центра ьные органы иммунитета: тимус и остный мозг;
2) защитная – на с изистой обо оч е минда ин и пейеровых б яше в ишечни е, в аппенди се распо ожены Т- имфоциты и В- имфоциты, изоцим и другие вещества, обеспечивающие защиту
На границе ротовой по ости и г от и в с изистой обо оч е распо агаются бо ьшие с оп ения |
имфоидной т ани. В |
||||
сово упности они |
образуют |
имфоэпите иа ьное г оточное |
о ьцо, о ружающее вход в дыхате ьные |
и |
|
пищеварите ьные пути. Наибо ее |
рупные с оп ения этого о ьца носят название минда ин. По месту их распо ожения |
||||
раз ичают небные минда ины, |
г оточную минда ину, язычную |
минда ину. Кроме перечис енных минда ин, |
в |
||
с изистой обо оч е переднего отде а пищеварите ьной труб и существует ряд с оп ений |
имфоидной т ани, |
из |
|||
оторых наибо ее |
рупными яв яются с оп ения в об асти с уховых труб – трубные минда ины и в же удоч е гортани |
||||
– гортанные минда ины. |
|
|
|
|
|
Миндалины выпо няют в организме важную защитную фун цию, обезвреживая ми робы, постоянно попадающие из внешней среды в организм через носовые и ротовое отверстия. Наряду с другими органами, содержащими имфоидную т ань, они обеспечивают образование имфоцитов, участвующих в реа циях гумора ьного и еточного иммунитета.
Развитие. Небные минда ины за адываются на 9-й неде е эмбриогенеза в виде уг уб ения псевдомногос ойного
реснитчатого эпите |
ия атера ьной стен и г от и, под оторым ежат омпа тно распо оженные мезенхимные ет и |
и многочис енные |
ровеносные сосуды. На 11—12-й неде е формируется тонзи ярный синус, эпите ий оторого |
перестраивается в многос ойный п ос ий, а из мезенхимы дифференцируется рети у ярная т ань; появ яются сосуды, в том чис е пост апи ярные вену ы с высо ими эндоте иоцитами. Происходит засе ение органа имфоцитами. На 14-й неде е среди имфоцитов опреде яются г авным образом Т- имфоциты (21 %) и немного В- имфоцитов (1 %). На 17— 18-й неде е появ яются первые имфатичес ие узе и. К 19-й неде е содержание Т- имфоцитов возрастает до 60 %, а В- имфоцитов — до 3 %. Рост эпите ия сопровождается формированием в эпите иа ьных тяжах пробо из ороговевающих ето .
Г оточная минда ина развивается на 4-м месяце внутриутробного периода из эпите ия и под ежащей мезенхимы дорса ьной стен и г от и. У эмбриона она по рыта многорядным мерцате ьным эпите ием. Язычная минда ина за адывается на 5-м месяце.
Минда ины достигают ма сима ьного развития в детс ом возрасте. Нача о инво юции минда ин совпадает с периодом по ового созревания.
Небные минда ины во взрос ом организме представ ены двумя те ами ова ьной формы, распо оженными по обеим сторонам г от и между небными дуж ами. Каждая минда ина состоит из нес о ь их с адо с изистой обо оч и, в собственной п астин е оторой распо ожены многочис енные имфатичес ие узе и (noduli lymphathici). От поверхности минда ины в г убь органа отходят 10–20 рипт (criptae tonsillares), оторые разветв яются и образуют вторичные рипты. С изистая обо оч а по рыта многос ойным п ос им неороговевающим эпите ием. Во многих местах, особенно в риптах, эпите ий часто бывает инфи ьтрирован (засе ен) имфоцитами и грану оцитами. Лей оциты, прони ающие в то щу эпите ия, обычно в бо ьшем и и меньшем о ичестве выходят на его поверхность и мигрируют навстречу ба териям, попадающим в по ость рта вместе с пищей и воздухом. Ми робы в минда ине а тивно
фагоцитируются ей оцитами и ма рофагами, при |
этом часть ей оцитов погибает. Под в иянием ми робов и |
|
раз ичных ферментов, выде яемых |
ей оцитами, |
эпите ий минда ины часто бывает разрушен. Одна о через |
не оторое время за счет размножения |
ето эпите иа ьного п аста эти участ и восстанав иваются. |
|
Собственная п астин а с изистой обо оч и образует небо ьшие сосоч и, вдающиеся в эпите ий. В рых ой во о нистой
соедините ьной т ани этого с оя распо ожены многочис енные имфатичес ие узе |
и. В центрах не оторых узе ов |
|
хорошо выражены бо ее свет ые участ и – герминативные центры. Лимфоидные узе |
и минда ин чаще всего отде ены |
|
друг от друга тон ими прос ой ами соедините ьной т ани. Одна о не оторые узе и могут с иваться. |
Мышечная |
|
п астин а с изистой обо оч и не выражена. |
|
|
Подс изистая основа, распо агающаяся под с оп ением имфоидных узе ов, образует во руг минда ины |
апсу у, от |
|
оторой в г убь минда ины отходят соедините ьнот анные перегород и. В этом с ое сосредоточены основные ровеносные и имфатичес ие сосуды минда ины и ветви язы ог оточного нерва, осуществ яющие ее иннервацию. Здесь же находятся и се реторные отде ы небо ьших с юнных же ез. Прото и этих же ез от рываются на поверхности с изистой обо оч и, распо оженной во руг минда ины. Снаружи от подс изистой основы ежат поперечнопо осатые мышцы г от и – ана ог мышечной обо оч и.

-лимфоциты (B- ет и, от bursa fabricii птиц, где впервые бы и обнаружены) — фун циона ьный тип имфоцитов, играющих важную ро ь в обеспечении гумора ьного иммунитета. При онта те с антигеном и и стиму яции со стороны
T- |
ето |
не оторые B- имфоциты трансформируются в п азматичес ие |
ет и, способные |
проду ции антите . Другие |
|||||||
а тивированные B- имфоциты превращаются в B- |
ет и памяти. Помимо проду ции антите , В- ет и выпо няют |
||||||||||
множество других фун ций: выступают в |
ачестве антигенпрезентирующих |
ето , |
продуцируют цито ины и |
||||||||
э зосомы*1+. |
|
|
|
|
|
|
|
|
|
||
Выде яют две субпопу яции В- |
ето : В-1 и B-2. Субпопу яцию В-2 состав яют обычные В- имфоциты, |
оторым |
|||||||||
относится всё с азанное выше. В-1 — это относите ьно небо ьшая группа В- |
ето , обнаруживаемая у че ове а и |
||||||||||
мышей. Они могут состав ять |
о о о 5% |
от общей попу яции B- |
ето . Та ие |
ет и появ яются в |
течение |
||||||
эмбриона ьного периода. На своей поверхности |
они э спрессируют |
IgM |
и небо ьшое |
о ичество (и и |
вовсе не |
||||||
э спрессируют) IgD. Мар ером этих ето яв яется CD5. Одна о он не яв яется обязате ьным омпонентом |
еточной |
||||||||||
поверхности. В эмбриона ьном периоде В1- |
ет и появ яются из ство овых |
ето |
остного мозга. В течение жизни пу |
||||||||
B-1- |
имфоцитов поддерживается за счёт а тивности специа изированных |
ето –предшественни ов и не попо няется |
|||||||||
за счёт |
ето , происходящих из |
остного мозга. К ет а–предшественница отсе яется из |
|
роветворной т ани на свою |
|||||||
анатомичес ую нишу — в брюшную и п евра ьную по ости — ещё в эмбриона ьном периоде. Ита , место обитания B-1- имфоцитов — прибарьерные по ости.
B-1- имфоциты значите ьно от ичаются от B-2- имфоцитов по антигенной специфичности продуцируемых антите . Антите а, синтезированные B-1- имфоцитами, не имеют значите ьного разнообразия вариабе ьных участ ов мо е у иммуног обу инов, но, напротив, ограничены в репертуаре распознаваемых антигенов, и эти антигены — наибо ее
распространённые |
соединения |
еточных |
стено ба терий. Все |
B-1- |
имфоциты — а |
бы один не с иш ом |
|||||||
специа изированный, но опреде ённо ориентированный (антиба териа ьный) |
он. Антите а, продуцируемые B-1- |
||||||||||||
имфоцитами, почти |
ис ючите ьно |
IgM, |
пере |
ючение |
ассов |
иммуног обу инов |
в B-1- |
имфоцитах не |
|||||
«предусмотрено». Та им образом, B-1- |
имфоциты — «отряд» противоба териа ьных «погранични ов» в прибарьерных |
||||||||||||
по остях, предназначенных д я |
быстрой |
реа ции на |
«просачивающиеся» |
через |
барьеры |
инфе ционные |
|||||||
ми роорганизмы из чис а широ о распространённых. В сыворот е |
рови здорового че ове а преоб адающая часть |
||||||||||||
иммуног обу инов |
— |
проду т |
синтеза |
а |
раз |
B-1- имфоцитов, |
т.е. это |
относите ьно по испецифичные |
|||||
иммуног обу ины антиба териа ьного назначения. |
|
|
|
|
|
|
|
||||||
ЭНДОКРИННАЯ СИСТЕМА
1. Классификация органов эндокринной системы. Гипоталамус. Источники развития. Нейросекреторные крупноклеточные и мелкоклеточные ядра. Особенности строения нейросекреторных клеток. Аксовазальные синапсы. Гормоны гипоталамуса, место и роль в эндокринной системе. Связь гипоталамуса с адено- и нейрогипофизом. Понятие об обратных связях.
Ле ция №16. Эндо ринная система. Центра ьное звено.
2. Классификация органов эндокринной системы. Гипофиз. Источники развития, строение, тканевой и клеточный состав адено- и нейрогипофиза, их функциональная характеристика. Морфо-функциональная характеристика аденоцитов. Связь гипофиза с гипоталамусом. Понятие об обратных связях.
Ле ция №16. Эндо ринная система. Центра ьное звено.
3. Классификация органов эндокринной системы. Эпифиз: источники развития, строение, тканевой и клеточный состав, функции. Место и роль эпифиза в эндокринной системе. Возрастные изменения.
Ле ция №16. Эндо ринная система. Центра ьное звено.
4. Классификация органов эндокринной системы. Щитовидная железа. Источники развития. Тканевой и клеточный состав. Особенности секреторного процесса в тироцитах, его регуляция. Роль гормонов тироцитов. Регенерация. Возрастные изменения. * 5. Структурно-функциональные особенности щитовидной железы новорожденного.
Ле ция №16. Эндо ринная система. Центра ьное звено.
Щитовидная железа окружена соединительнотканной капсулой, прослойки которой направляются вглубь и разделяют орган на дольки. В этих прослойках располагаются многочисленные сосуды микроциркуляторного русла и нервы.
Основными структурными компонентами паренхимы железы являются фолликулы - замкнутые шаровидные или слегка вытянутые образования с полостью внутри. Стенка фолликулов образована одним слоем эпителиальных клеток - фолликулярных тироцитов, среди которых встречаются одиночные клетки нейрального происхождения - парафолликулярные С-клетки.
Вдольках щитовидной железы можно выделить фолликулярные комплексы, или микродольки, которые состоят из группы фолликулов, окруженных тонкой соединительнотканной капсулой.
Впросвете фолликулов накапливается коллоид - секреторный продукт тироцитов, представляющий собой вязкую жидкость, состоящую в основном из тироглобулина. Размер фолликулов и образующих их тироцитов варьирует в нормальных физиологических условиях. В небольших формирующихся фолликулах, еще не

заполненных коллоидом, эпителий однослойный призматический. По мере накопления коллоида размеры фолликулов увеличиваются, эпителий становится кубическим, а в сильно растянутых фолликулах, заполненных коллоидом эпителий становится плоским. Основная масса фолликулов в норме образована тироцитами кубической формы. Увеличение размеров фолликулов обусловлено пролиферацией, ростом и дифференцировкой тироцитов, сопровождаемой накоплением коллоида в полости фолликула.
Фолликулы разделяются тонкими прослойками рыхлой волокнистой соединительной ткани с многочисленными кровеносными и лимфатическими капиллярами, оплетающими фолликулы, а также тучными клетками и лимфоцитами.
Фолликулярные эндокриноциты, или тироциты, - это железистые клетки, составляющие большую часть стенки фолликулов. В фолликулах тироциты располагаются в один слой на базальной мембране.
Тироциты изменяют свою форму от плоской до циллиндрической в зависимости от функционального состояния железы. При умеренной функциональной активности щитовидной железы тироциты имеют кубическую форму и шаровидные ядра. Коллоид, секретируемый ими, заполняет в виде гомогенной массы просвет фолликула. На апикальной поверхности тироцитов, обращенной к просвету фолликула, имеются микроворсинки. По мере усиления тироидной активности количество и размеры микроворсинок возрастают. Базальная поверхность тироцитов, обращенная к поверхности фолликула, почти гладкая. Соседние тироциты тесно связаны между собой многочисленными десмосомами и хорошо развитыми терминальными пластинками. По мере возрастания тироидной активности на боковых поверхностях тироцитов возникают пальцевидные выступы (или интердигитации), входящие в соответствующие вдавления боковой поверхности соседних клеток.
Функция тироцитов заключается в синтезе и выделении йод-содержащих тиреоидных гормонов - Т3, или трийодтиронина, и Т4, или тироксина.
В тироцитах хорошо развиты органеллы, особенно участвующие в белковом синтезе. Белковые продукты, синтезируемые тироцитами, выделяются в полость фолликула, где завершается образование йодированных тирозинов и тиронинов (т.е. аминокислот, входящих в состав крупной и сложной молекулы тироглобулина). Тироидные гормоны могут попасть в циркуляцию лишь после высвобождения из этой молекулы (т.е. после расщепления тироглобулина).
Когда потребности организма в тироидном гормоне возрастают и функциональная активность щитовидной железы усиливается, тироциты фолликулов принимают призматическую форму. Интрафолликулярный коллоид при этом становится более жидким и пронизывается многочисленными ресорбционными вакуолями.
Ослабление функциональной активности (гипофункция) щитовидной железы проявляется, наоборот, уплотнением коллоида, его застоем внутри фолликулов, диаметр и объем которых значительно увеличиваются; высота тироцитов уменьшается, они принимают уплощенную форму, а их ядра вытягиваются параллельно поверхности фолликула.
В секреторном цикле фолликулярных эндокриноцитов различают две основные фазы: фазу продукции и фазу выведения гормонов.
Фаза продукции включает:
поступление предшественников тироглобулина (аминокислот, углеводов, ионов, воды, йодидов), приносимых из кровеносного русла в тироциты;
синтез фермента тиропероксидазы, окисляющей йодиды и обеспечивающей их соединение с тироглобулином на поверхности тироцитов и в полости фолликула и образование коллоида;
синтез полипептидных цепочек самого тироглобулина в гранулярной эндоплазматической сети и их
гликозилирование (т.е. соединение с нейтральными сахарами и сиаловой кислотой) с помощью тиропероксидазы (в аппарате Гольджи).
Фаза выведения включает резорбцию тироглобулина из коллоида путем пиноцитоза и его гидролиз с помощью лизосомных протеаз с образованием гормонов тироксина и трийодтиронина, а также выведение этих гормонов через базальную мембрану в гемокапилляры и лимфокапилляры.
Гипофизарный тиротропный гормон (ТТГ) усиливает функцию щитовидной железы, стимулируя поглощение тироглобулина микроворсинками тироцитов, а также его расщепление в фаголизосомах с высвобождением активных гормонов.
Тиреоидные гормоны (Т3 и Т4) участвуют в регуляции метаболических реакций, влияют на рост и дифференцировку тканей, особенно на развитие нервной системы.
Второй вид эндокриноцитов щитовидной железы - парафолликулярные клетки, или C-клетки, или же кальцитониноциты. Это клетки нейрального происхождения. Их главная функция - выработка тиреокальцитонина, снижающего уровень кальция в крови.
Во взрослом организме парафолликулярные клетки локализуются в стенке фолликулов, залегая между основаниями соседних тироцитов, но не достигают своей верхушкой просвета фолликула. Кроме того, парафолликулярные клетки располагаются также в межфолликулярных прослойках соединительной ткани. По размерам парафолликулярные клетки крупнее тироцитов, имеют округлую, иногда угловатую форму. Парафолликулярные клетки осуществляют биосинтез пептидных гормонов -кальцитонина и соматостатина, а также участвуют в образовании нейроаминов (норадреналина и серотонина) путем декарбоксилирования соответствующих аминокислот-предшественников.
Секреторные гранулы, заполняющие цитоплазму парафолликулярных клеток, обнаруживают сильную осмиофилию и аргирофилию (т.е. эти клетки хорошо выявляются при импрегнации солями осмия и серебра).
Васкуляризация. Щитовидная железа обильно снабжается кровью. За единицу времени через щитовидную железу проходит приблизительно столько же крови, сколько через почки, причем интенсивность кровоснабжения значительно увеличивается при усилении функциональной активности органа.

Иннервация. В щитовидной железе много симпатических и парасимпатических нервных волокон. Стимуляция адренергических нервных волокон приводит к небольшому усилению, а парасимпатических - к угнетению функции фолликулярных эндокриноцитов. Основная же регулирующая роль принадлежит тиротропному гормону гипофиза. Парафолликулярные клетки невосприимчивы к тиротропному гормону, но отчетливо реагируют на активирующие симпатические и угнетающие парасимпатические нервные импульсы.
Регенерация щитовидной железы в физиологических условиях осуществляется очень медленно, однако способность паренхимы к пролиферации велика. Источником роста тироидной паренхимы является эпителий фолликулов. Нарушение механизмов регенерации может приводить к разрастанию железы с образованием зоба.
У новорожденного масса щитовидной же езы |
о еб ется от 1 до 5 г, диаметр фо |
и у а состав яет 60-70 м м, |
в |
||||||||
возрасте одного года - 100 м м, 3 |
ет - 120-150 м м, 6 ет - 200 м м, 12-15 |
ет - 250 м м. |
Уве ичение |
о ичества |
|||||||
фо и у ов происходит путем их поч ования. За счет де ения |
ето |
стен а фо |
и у а уто щается в |
а ом-то ее |
|||||||
участ е, в месте уто щения образуется по ость, и сформированный |
та им |
образом новый дочерний фо и у |
|||||||||
отде яется от основного. Пос е отде ения фо |
и у |
по рывается соедините ьнот анной |
апсу ой. Фо |
и у ярный |
|||||||
эпите ий щитовидной же езы у новорожденных |
убичес ий и и ци индричес ий. По |
мере роста организма он |
|||||||||
заменяется на убичес ий и п ос ий, |
оторый хара терен д я фо |
и у ов щитовидной же езы взрос ого че ове а. |
|||||||||
Уже на втором-третьем месяцах внутриутробного развития в центре фо |
и у а появ яется |
о оид. На 14-15-й неде е |
|||||||||
щитовидная же еза начинает прояв ять свою се реторную а тивность. Способность |
онцентрации йода в щитовидной |
||||||||||
же езе появ яется на 14-й неде е, а связывание его в органичес ую форму отмечается от 15-й до 19-й неде и эмбриона ьного развития. К моменту рождения щитовидная же еза яв яется фун циона ьно а тивной, одна о и в постната ьном развитии в соответствии с продо жающимся морфо огичес им созреванием происходит совершенствование ее фун ции.
5. Классификация органов эндокринной системы. Околощитовидные железы. Источники развития. Тканевой и клеточный состав. Секреторная функция железы и механизмы ее регуляции. Возрастные изменения.
Ле ция №16. Эндо ринная система. Центра ьное звено.
Околощитовидные (паращитовидные) железы
Околощитовидные железы (обычно в количестве четырех) расположены на задней поверхности щитовидной железы и отделены от нее капсулой.
Функциональное значение околощитовидных желез заключается в регуляцииметаболизма кальция. Они вырабатывают белковый гормон паратирин, или паратгормон, который стимулирует резорбцию кости остеокластами, повышая уровень кальция в крови. Сами остеокласты не имеют рецепторов к паратгормону, - его действие опосредовано другими клетками костной ткани - остеобластами.
Кроме этого паратгормон уменьшает выведение кальция почками, а также усиливает синтез метаболита витамина D, который, в свою очередь, повышает всасывание кальция в кишечнике.
Развитие. Околощитовидные железы закладываются у зародыша как выступы из эпителия III-ей и IV-ой пар жаберных карманов глоточной кишки. Эти выступы отшнуровываются, и каждый из них развивается в отдельную околощитовидную железу, причем из IV пары жаберных карманов развивается верхняя пара желез, а из III пары развивается нижняя пара околощитовидных желез, а также вилочковая железа - тимус.
III.Строение околощитовидной железы
Каждая околощитовидная железа окружена тонкой соединительнотканной капсулой. Ее паренхима представлена трабекулами - эпителиальными тяжами эндокринных клеток - паратироцитов. Трабекулы разделены тонкими прослойками рыхлой соединительной ткани с многочисленными капиллярами. Хотя между паратироцитами хорошо развиты межклеточные щели, соседние клетки связаны интердигитациями и десмосомами. Различают два типа клеток: главные паратироциты и оксифильные паратироциты.
Главные клетки секретируют паратирин, они преобладают в паренхиме железы, имеют небольшие размеры и полигональную форму. В периферических зонах цитоплазма базофильна, где рассеяны скопления свободных рибосом и секреторные гранулы. При усилении секреторной активности паращитовидных желез главные клетки увеличиваются в объеме. Среди главных паратироцитов также различают два типа: светлые и темные. В цитоплазме светлых клеток встречаются включения гликогена. Считают, что светлые клетки - это неактивные, а темные клетки - функционально активные паратироциты. Главные клетки осуществляют биосинтез и выделение паратгормона.
Второй тип клеток - оксифильные паратироциты. Они малочисленны, располагаются поодиночке или группами. Они значительно крупнее, чем главные паратироциты. В цитоплазме видны оксифильные гранулы, огромное количество митохондрий при слабом развитии других органелл. Их рассматривают как стареющие формы главных клеток. У детей эти клетки единичны, с возрастом их число возрастает.
На секреторную активность околощитовидных желез не оказывают влияния гипофизарные гормоны. Околощитовидная железа по принципу обратной связи быстро реагирует на малейшие колебания в уровне кальция в крови. Ее деятельность усиливается при гипокальциемии и ослабляется при гиперкальциемии. Паратироциты обладают рецепторами, способными непосредственно воспринимать прямые влияния ионов кальция на них.