

Дегидрирование сукцината
Образование малата из фумарата
Дегидрирование малата

Механизмы регуляции скорости цикла лимонной кислоты.
Регуляция цитратного цикла.
В большинстве случаев скорость реакций в метаболических циклах определяется их начальными реакциями.
В ЦТК важнейшая регуляторная реакция - образование цитрата из оксалоацетата и ацетил-КоА, катализируемая цитратсинтазой.
Эта реакция ускоряется при повышении концентрации оксалоацетата - субстрата реакции
тормозится продуктом реакции - цитратом.
Когда отношение NADH/NAD+ снижается, скорость окисления маната в оксалоацетат возрастает.
Повышение концентрации оксалоацетата ускоряет цитратсинтазную реакцию.
Скорость реакции снижается при повышении концентрации АТФ, сукцинил-КоА и длинноцепочечных жирных кислот.
ПДК активируется пируватом, NAD+, КоA; ингибируется NADН и ацетил – КоА
Цитратсинтаза (реакция ускоряется при повышении концентрации оксалоацетата и замедляется при повышении концентрации цитрата, АТФ, NADH и сукцинил-КоA).
Изоцитратдегидрогеназа аллостериески активируется АДФ, ионами кальция, ингибируется NADH и сукцинил-КоA .
Альфа-кетоглутаратдегидрогеназный комплекс, имеющий сходное строение с пируватдегидрогеназным, в отличие от последнего, не имеет в своём составе регуляторных субъединиц. Главный механизм регуляции а-кетоглутаратдегидрогеназного комплекса - ингибированиереакции NADH и сукцинил-КоA.
а-Кетоглутаратдегидрогеназный комплекс, как и изоцитратдегидрогеназа, активируется Са+, а при повышении концентрации АТФ скорости обеих реакций снижаются.
14. Анаплеротические реакции цикла лимонной кислоты (уравнения реакций, ферменты, биологическая роль). Анаболическое значение цикла лимонной кислоты.
Анаболические функции цитратного цикла
Цикл лимонной кислоты - один из амфиболических путей метаболизма. В нём осуществляются не только окислительные превращения энергетических субстратов до конечных продуктов СО2 и Н2О, но и происходит образование субстратов для других метаболических путей (рис. 6-28).
Убыль промежуточных продуктов цикла восполняется в реакциях, катализируемых специфическими ферментами. В нормальных условиях реакции, отвлекающие промежуточные продукты из цикла и восполняющие их убыль, находятся в состоянии динамического равновесия, так что концентрация этих продуктов в митохондриях остаётся постоянной.
Реакции, обеспечивающие пополнение фонда промежуточных продуктов ЦТК, называются анаплеротическими (пополняющими).
Важнейшая из них - реакция синтеза оксалоацетата из пирувата. Эту реакцию катализирует митохондриальный фермент - пируваткарбоксилаза.
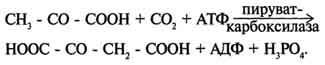
Пируваткарбоксилаза - сложный олигомерный фермент. Молекула фермента содержит 4 простетические группы, представленные биотином), который ковалентно связан амидной связью с е-аминогруппами остатков лизина, находящегося в активном центре фермента (рис. 6-29).
Пируваткарбоксилаза - регуляторный фермент. Если концентрация ацетил-КоА увеличивается, то он действует как аллостерический активатор пируваткарбоксилазы, ускоряя образование оксалоацетата. Таким образом, избыток ацетил-КоА способствует активации цитратного цикла.
Если для цикла лимонной кислоты не хватает оксалоацетата или какого-нибудь другого промежуточного продукта, то карбоксилирование пирувата ускоряется. В этой реакции в качестве источника энергии используется АТФ.
Реакция протекает в 2 стадии.
На первой стадии происходит активация СО2 путём присоединения к одному из атомов азота в молекуле биотина. Эта реакция сопряжена с гидролизом АТФ.
АТФ + СО2+ Е-биотин + Н2О → АДФ + Н3РО4 + Е-биотин-СОО- + 2 Н+.
На второй стадии активированная карбоксильная группа переносится на пируват.
Е-биотин-СОО" + Пируват -" Е-биотин + Оксалоацетат.
Метаболиты цитратного цикла используются не только как субстраты синтеза углеродного скелета ряда соединений, но и являются донорами водорода для образования восстановленных коферментов, участвующих в реакциях синтеза жирных кислот, стероидов и других веществ (см. разделы 8, 10, 11). Два метаболита нитратного цикла могут дегидрироваться при участии NADP-зависимых дегидрогеназ: малата и изоцитрата.
Например, малат может поступать из митохондрий в цитозоль клетки. В цитозоле находится NADP-зависимая дегидрогеназа (малик-фермент), катализирующая реакцию:

+Малат и изоцитрат обеспечивают образование около половины общего фонда NADPH, используемого в восстановительных синтезах; вторая половина образуется в пентозофосфатном пути превращения глюкозы.
