

Разобщение тканевого дыхания и окислительного фосфорилирования. Терморегуляторная функция тканевого дыхания. Термогенная функция энергетического обмена в бурой жировой ткани.
Эффективность окислительного фосфорилирования в митохондриях определяется как отношение величины образовавшегося АТФ к поглощенному кислороду: АТФ/О или Р/О (коэффициент фосфорилирования). Экспериментально определяемые значения Р/О, как правило, оказываются меньше 3, а учитывая необходимость определенных затрат протонного градиента на перенос АТФ в цитоплазму с обменом ее на АДФ и Н2РО4─, требующим одновременного переноса Н+ в митохондрию, полагают, что на 1 моль О, или на два моля электронов, использованных для восстановления кислорода до воды в митохондриях, в клетке не может образоваться более 2,5 моль АТФ. Это свидетельствует о том, что процесс дыхания не полностью сопряжен с фосфорилированием. При полном сопряжении этих процессов 220 кдж энергии, освобождающейся при окислении 1 моль НАДН кислородом до воды и НАД+, хватило бы на образование более 5 моль АТФ. Степень сопряжения зависит главным образом от самого механизма сопряжения, при котором энергия электронов расходуется на выкачивание протонов из митохондрий и создание протонного градиента (электрохимического мембранного потенциала), который в свою очередь используется для синтеза АТФ. В этом процессе важнейшую роль играет целостность митохондриальной мембраны, удерживающей электрохимический потенциал, создаваемый транспортом электронов.
Некоторые химические вещества, ионофоры, могут, растворяясь в липидном бислое митохондриальной мембраны, переносить ионы через мембрану; протонофоры могут усиливать протонную проводимость митохондриальной мембраны, перенося протоны из межмембранного пространства через мембрану в матрикс, минуя протонные каналы АТФ-синтазы, а другие ионофоры переносят другие ионы. В результате таких процессов уменьшается электрохимический мембранный потенциал и уменьшается, и даже прекращается синтез АТФ. Это явление называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а
+
концентрация АДФ увеличивается. При этом возрастает скорость окисления ФАДН2 и НАДН, а также количество потребленного кислорода, но энергия в основном выделяется в виде теплоты. Происходит рассеивание энергии, которая не фиксируется в виде макроэргической связи молекулы АТФ, а принимает вид тепловой энергии. Соединения, обеспечивающие протонную проводимость и понижающие электрохимический потенциал, называются раз-
общителями.
Одно из таких веществ 2,4-динитрофенол, легко переходящий из ионизированной формы в неионизированную, присоединяя протон в межмембранном пространстве и перенося его в матрикс. Примерами разобщителей могут также быть лекарства (дикумарол) или метаболиты, которые образуются в организме (билирубин), а также тироксин – гормон щитовидной железы, но эти вещества проявляют свое разобщающее действие только при высокой концентрации.
Разобщение окисления и фосфорилирования также может возникать при некоторых воздействиях на организм или ткани (например, при переохлаждении)
Терморегуляторная функция ЦПЭ
На синтез молекул АТФ расходуется примерно 40-45% всей энергии электронов, переносимых по ЦПЭ, приблизительно 25% тратится на работу по переносу веществ через мембрану. Остальная часть энергии рассеивается в виде теплоты и используется теплокровными животными на поддержание температуры тела. Кроме того, дополнительное образование теплоты может происходить при разобщении дыхания и фосфорилирования. Разобщение окислительного фосфорилирования может быть биологически полезным. Оно позволяет генерировать тепло для поддержания температуры тела у новорождённых, у зимнес-пящих животных и у всех млекопитающих в процессе адаптации к холоду. У новорождённых, а также зимнеспящих животных существует особая ткань, специализирующаяся на теплопродукции посредством разобщения дыхания и фосфорилирования - бурый жир. Бурый жир содержит много митохондрий. В мембране митохондрий имеется большой избыток дыхательных ферментов по сравнению с АТФ-синтазой. Около 10% всех белков приходится на так называемый разобщающий белок (РБ-1) - термогенин. Бурый жир имеется у новорождённых, но его практически нет у взрослого человека. В последние годы появились факты, свидетельствующие о существовании в митохондриях разных органов и тканей млекопитающих разобщающих белков, похожих по своей структуре на РБ-1 бурой жировой ткани. По своей структуре термогенин близок к АТФ/АДФ-антипортеру, но не способен к транспорту нуклеотидов, хотя сохранил способность переносить анионы жирных кислот, служащих разобщителями.
На внешней стороне мембраны анион жирной кислоты присоединяет протон и в таком виде пересекает мембрану; на внутренней стороне мембраны диссоциирует, отдавая протон в матрикс и тем самым снижает протонный градиент. Образующийся анион возвращается на наружную сторону мембраны с помощью АТФ/ АДФ-антипортера.
При охлаждении стимулируется освобождение норадреналина из окончаний симпатических нервов. В результате происходят активация липазы в жировой ткани и мобилизация жира из жировых депо. Образующиеся свободные жирные кислоты служат не только "топливом", но и важнейшим регулятором разобщения дыхания и фосфорилирования.
Бурая жировая ткань
При фосфорилировании АТФ-синтазой АДФ до АТФ используется не вся энергия протонного градиента, лишь его часть тратится на совершение работы (синтез АТФ), а другая часть рассеивается в виде тепла.

Особые клетки организма умеют увеличивать долю рассеиваемой тепловой энергии – это клетки бурой жировой ткани. В отличие от белых жировых клеток бурые адипоциты содержат большое количество митохондрий, которые и придают им буро-красный цвет. Во внутренней мембране митохондрий этих клеток имеется белок термогенин (до 15% от всех белков митохондрий), относящийся к семейству разобщающих белков, называемых UCP-белки (англ. uncoupling protein).
При охлаждении организма бурые адипоциты получают сигналы по симпатическим нервам, и в них активируется расщепление жира – липолиз. Окисление жиров приводит к получению большого количества НАДН и ФАДН2, активизации работы дыхательной цепи и возрастанию электрохимического градиента. Однако АТФ-синтазы в мембранах митохондрий этих клеток относительно мало, зато много термогенина, близкого по строению к Fо-субъединице АТФ-синтазы. Термогенин является каналом во внутренней мембране через который в матрикс проходит часть ионов Н+ и снижается протонный градиент. В итоге АТФ не синтезируется, несмотря на непрерывно возрастающий (по правилу дыхательного контроля) катаболизм. Благодаря термогенину большая часть энергии ионов водорода рассеивается в виде тепла, подогревая протекающую через ткань кровь и обеспечивая поддержание температуры тела при охлаждении.
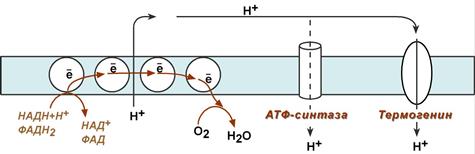
Роль термогенина в снижении протонного градиента
Открыто несколько типов UCP-белков: UCP-1 (термогенин) преобладает в бурой жировой ткани, UCP-2 – есть и в бурой и белой жировой ткани, UCP-3 – в cкелетных мышцах, UCP-4 и UCP-5 – обнаружены в нейронах.
Бурая жировая ткань широко представлена в верхней части спины у животных, впадающих в зимний сон (медведи), у детенышей животных и человеческих младенцев. Отдельные скопления бурых жировых клеток есть и у взрослого человека, они расположены в верхней части спины, между лопаток, под мышками, по ходу крупных кровеносных сосудов.
В настоящее время постулировано существование бежевой жировой ткани, количество которой увеличивается при влиянии ряда факторов. Она тоже отвечает за образование тепла.
9. Катаболизм основных пищевых веществ (углеводы, жиры, аминокислоты и белки). Понятие об общем и специфических путях катаболизма. Общий путь катаболизма: окисление пирувата и ацетил-КоА. Биологическое значение, локализация в клетке.
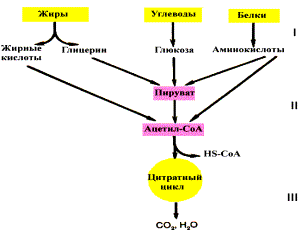
Если процесс катаболизма рассматривать с общей точки зрения, то можно выделить три основные его части:
Расщепление в пищеварительном тракте. Это гидролитические реакции, превращающие сложные пищевые вещества в относительно небольшое число простых метаболитов: глюкоза, аминокислоты, глицерин, жирные кислоты.
Специфические пути катаболизма. Простые метаболиты подвергаются специфическим реакциям расщепления, в результате которых образуется либо пировиноградная кислота, либо ацетил — КоА. Причем ацетил — КоА может образоваться из пирувата в результате окислительного декарбоксилирования. Могут также образоваться другие соединения, непосредственно включающиеся в цитратный цикл.
Цитратный цикл и дыхательная цепь завершают расщепление пищевых веществ до конечных продуктов — СО2 и Н2О.
Следовательно, начиная со стадии образования пирувата происходит унификация путей катаболизма. Из большого числа исходных соединений образуется всего два — пируват и ацетил — СоА. Процесс, начинающийся от пирувата, называется общим путем катаболизма и в свою очередь включает:
окислительное декарбоксилирование пирувата
цитратный цикл.
Именно в общем пути катаболизма образуется основная масса субстратов для реакций дегидрирования. Совместно с дыхательной цепью и окислител
Пируват окисляется до уксусной кислоты
Пировиноградная кислота (ПВК, пируват) является продуктом окисления глюкозы и некоторых аминокислот. Ее судьба различна в зависимости от доступности кислорода в клетке. В анаэробных условиях она восстанавливается до молочной кислоты. В аэробных условиях пируват симпортом с ионами Н+, движущимися по протонному градиенту, проникает в митохондрии. Здесь происходит его превращение до уксусной кислоты, переносчиком которой служит коэнзим А.