

Процессы катаболизма и анаболизма в клетках. Эндергонические и экзергонические реакции в живой клетке. Макроэргические соединения: определение, примеры.
Эндергонические и экзергонические реакции в живой клетке

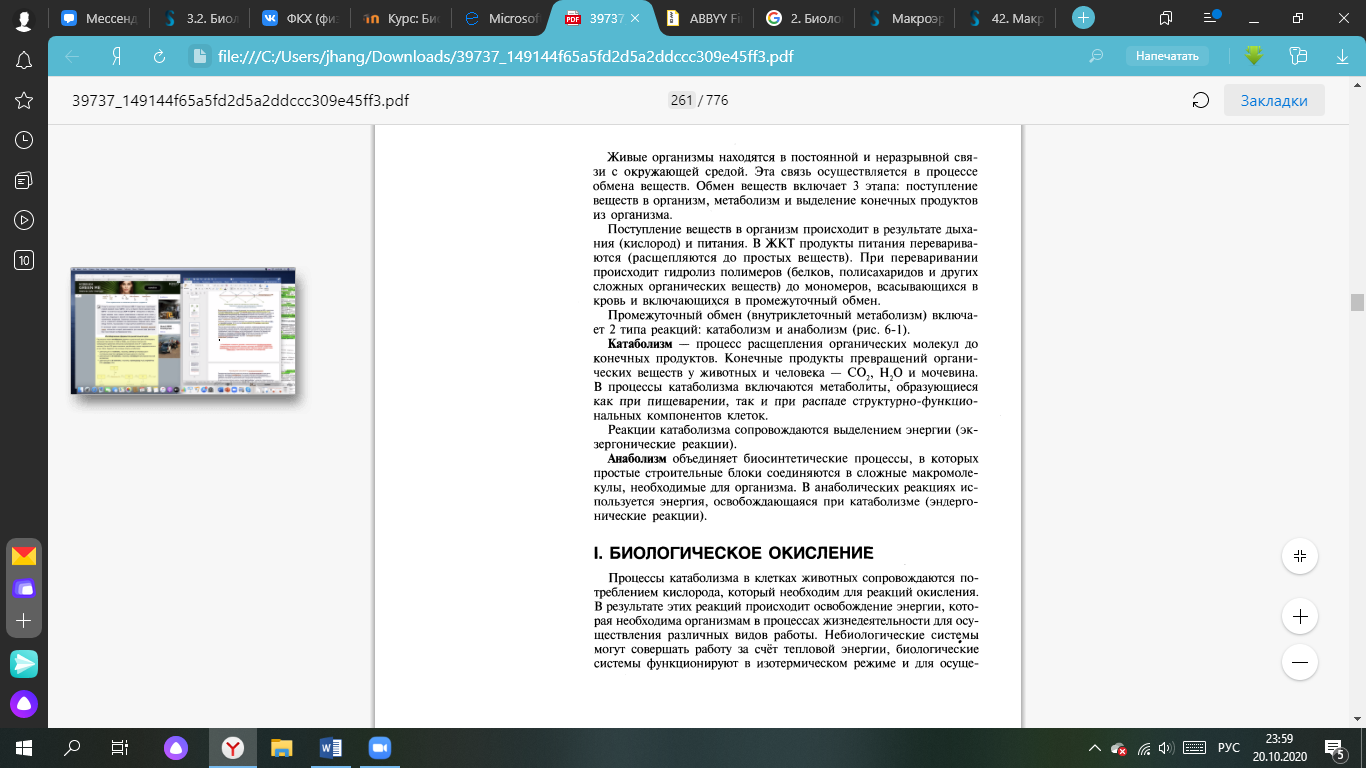
В биологических системах термодинамически невыгодные (эндергонические) реакции могут протекать лишь за счёт энергии экзергонических реакций. Такие реакции называют энергетически сопряжёнными. Многие из этих реакций происходят при участии аденозинтрифосфата (АТФ), играющего роль сопрягающего фактора.
Макроэргические соединения: определение, примеры.
Макроэргические соединения– это вещества, содержащие богатые энергией связи.
К макроэргическим соединениям относятся:
Макроэргические соединения классифицируются по типу связи, несущей дополнительную энергию:
1. Фосфоангидридная связь. Такую связь имеют все нуклеотиды: нуклеозидтрифосфаты (АТФ, ГТФ, ЦТФ, УТФ, ТТФ) и нуклеозиддифосфаты (АДФ, ГДФ, ЦДФ, УДФ, ТДФ).
2. Тиоэфирная связь. Примером являются ацил-производные коэнзима А: ацетил-SКоА, сукцинил-SКоА, и другие соединения любой жирной кислоты c HS-КоА.
3. Гуанидинфосфатная связь – присутствует в креатинфосфате, запасном макроэрге мышечной и нервной ткани.
4. Ацилфосфатная связь. К таким макроэргам относится метаболит гликолиза 1,3-дифосфоглицериновая кислота (1,3-дифосфоглицерат). Она обеспечивает синтез АТФ в реакции субстратного фосфорилирования.
5. Енолфосфатная связь. Представитель – фосфоенолпируват, метаболит гликолиза. Он также обеспечивает синтез АТФ в реакции субстратного фосфорилирования в гликолизе.
Самое важное макроэргическое соединение — АТФ. Используя энергию, заключенную в макроэргических связях АТФ, при действии ферментов, переносящих фосфатные группы, можно получить другие макроэргические соединения, например, ГТФ (гуанозинтрифосфорная кислота), ФЕП (фосфоенолпировиноградная кислота) и др.
Образуется АТФ в процессах биологического окисления и при фотосинтезе. Энергия макроэргических связей используется для совершения любой работы: активации соединений (например, глюкозы, чтобы могла начаться цепь ее окислительных превращений), синтеза биополимеров (нуклеиновых кислот, белков, полисахаридов), избирательного поглощения веществ из окружающей клетку среды и выброса из клетки ненужных продуктов, мышечного сокращения и восстановления активного состояния организма и т. д. Запас этих соединений позволяет организму быстро реагировать на изменение внешних условий и совершать физическую работу.
При спортивной тренировке содержание макроэргических соединений в мышцах и скорость их образования возрастают.
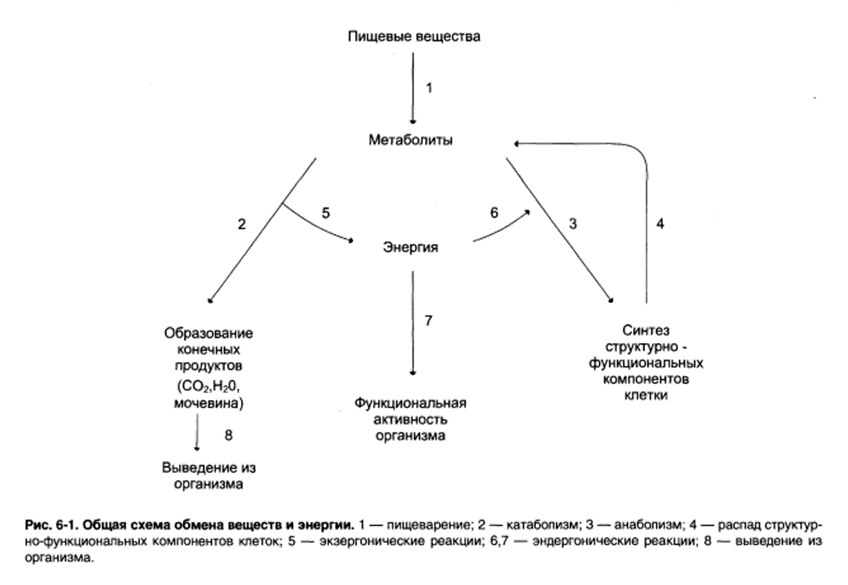
2. Биологическое окисление. Биологические функции биологического окисления в клетке. Дегидрирование субстратов и восстановление кислорода как источник энергии для синтеза АТФ. Виды фосфорилирования как реакции образования АТФ.


Функции био.окисления
Дегидрирование субстратов и восстановление кислорода как источник энергии для синтеза АТФ
АТФ – чрезвычайно важная молекула клетки. Она непрерывно синтезируется и используется. Энергия для синтеза АТФ поступает от субстратов, отдающих электроны в ходе их дегидрирования. Электроны извлекаются из субстратов в ходе гликолиза и гликогенолиза (в цитозоле), при окислении жирных кислот и в общих путях метаболизма: при преобразовании пирувата в ацетил-КоА и в цикле трикарбоновых кислот (в митохондриальном матриксе). Молекулы НАДН и ФАДН2 переносят эти электроны в дыхательную цепь, локализованную во внутренней митохондриальной мембране. Энергия этих электронов используется для создания трансмембранного протонного градиента потенциала, используемого для синтеза АТФ.
Виды фосфорилирования как реакции образования АТФ: окислительное, субстратное, фотофосфорилирование.
окислительное фосфорилирование – образование АТФ за счет освобождения и аккумуляции энергии, выделяемой в процессе окисления питательных веществ. Этот механизм протекает в митохондриях и является основным путем образования АТФ.
субстратное фосфорилирование – энергия, необходимая для образования АТФ (ГТФ) высвобождается при гидролизе связей других макроэргических соединений. Синтез АТФ происходит за счет энергии макроэргических связей S. Механизм сопряжения не требует участие мембраны. К субстратам, богатым энергией, относятся фосфоглицериновая кислота, фосфоэнолпируват (ФЭП), сукцинил-СоА, креатинфосфат, и ряд других.
Фотофосфорили́рование — процесс синтеза АТФ из АДФ за счёт энергии света. Как и в случае окислительного фосфорилирования, энергия света расходуется на создание протонного градиента на мембране тилакоидов или клеточной мембране бактерии, который затем используется АТФ-синтазой. Есть у всех фототрофных эукариот, бактерий и архей.
Различают два типа фосфорилирования — циклическое, сопряжённое с циклическим потоком электронов в электрон-транспортной цепи, и нециклическое, сопряжённое с прямым потоком электронов от H2O к НАДФ+ в случае эукариот или другого донора электрона в случае бактерий, например, H2S.