

5.3 Цитокинины
В 1955 было выделено новое химическое вещество, являющееся по составу 6-фурфуриламинопурином, который образуется из дезоксиаденозина при определенных условиях деградации ДНК. Поскольку его добавление к питательной среде, содержащей сахарозу, элементы минерального питания, витамины и ауксин, вызывало переход клеток изолированной сердцевины стебля табака к делению, это вещество получило название кинетин (от слова кинез — деление).
Вскоре оно было синтезировано. Последовал синтез большого числа его химических аналогов, обладающих такой же или даже более высокой биологической активностью. Все эти вещества были объединены под общим названием цитокинины.
Первый из цитокининов — зеатин — 6-(4'-окси-3'-метил-транс-2'-бутенил-амино)пурин был выделен из семян кукурузы в стадии молочной спелости и идентифицирован Литамом в 1964 г. Это вещество содержится в растениях в очень малых количествах. Для получения 1 мг зеатина потребовалось переработать 70 кг семян кукурузы. При этом нужно иметь в виду, что развивающиеся семена отличаются наиболее высоким содержанием цитокининов.
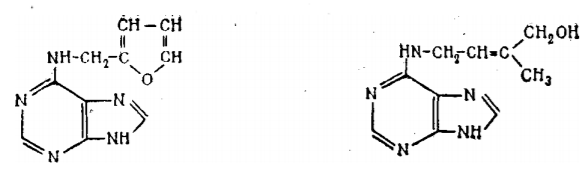
Рис. 6 — Кинетин (слева) и зеатин (справа)
Обязательным для физиологической активности этих соединений является присутствие в их молекуле радикала у аминогруппы при шестом атоме углерода пуринового кольца. Строение этого радикала также подчиняется следующему правилу: у соединений с алифатическим радикалом активность увеличивается по мере удлинения радикала от 1 до 6 атомов углерода и резко падает при дальнейшем его удлинении.
Введение в пуриновое кольцо дополнительных радикалов, как правило, снижает цитокининовую активность соединения. Однако известны случаи, когда введение метильной группы в 8 и 9 положение пуринового кольца повышало активность.[1]
Цитокинины участвуют во многих физиологических процессах растений, регулируют деления клеток, морфогенез побега и корня, созревание хлоропластов, линейный рост клетки, образование добавочных почек и старение.[9] Соотношение ауксинов к цитокининам является ключевым фактором деления клеток и дифференцировки тканей растения.
В то время как эффект цитокининов на сосудистые растения является плейотропным, цитокинины вызывают изменения роста протонемы у мхов. Образование почек можно считать вариантом дифференцировки клеток и этот процесс является очень специфическим эффектом цитокининов.[10]
Главные функции цитокининов — они стимулируют деление клеток и дифференцировку, усиливая синтез ДНК и белка, активируют растяжение клеток листьев у двудольных растений, образование клубней, снимают апикальное доминирование, вызывая заложение и рост пазушных почек. Цитокинины стимулируют растяжение у изолированных семядолей, отрезков стеблей или колеоптилей.
Цитокинины обладают высокой аттрагирующей способностью, т.е. притягивают ассимиляты (аминокислоты, углеводы) и регуляторные вещества к клеткам и тканям, например, к зоне деления клеток, содержащим большое количество этого гормона, что было доказано с помощью следующего опыта. На левую нижнюю часть листа табака нанесли раствор 14С-глюкозы, на правую верхнюю часть — раствор кинетина. Скоро радиоактивная метка была обнаружена в правой верхней части листа, следовательно, сюда поступила глюкоза.
Эти гормоны задерживают старение листьев, что легче всего можно увидеть в опытах с изолированными листьями. Когда отрезанный лист ставят в воду, старение наступает быстро: разрушается хлорофилл, лист желтеет, в нем усиливается распад белка, РНК. Однако старение листа задерживается, если на черешке образуются придаточные корни, синтезирующие цитокинины. Задержка старения связана еще и с тем, что обработка листа, например, кинетином вызывает образование в хлоропластах новых тилакоидов стромы и гран, разрушившихся при старении, усиливает синтез хлорофилла. Формирование хлоропластов у цветковых растений при добавлении экзогенного цитокинина происходит даже в темноте.[8]