

Справочный материал по Физиологии.
Глава 17– Интегративныефункциинервнойсистемы.
Под интегративными функциями понимают аналитико-синтетическую деятельность коры больших полушарий и многих подкорковых структур. Эти функции — обучение, память, рассудок, речь, сознание и некоторые другие — обычно называют высшими (или когнитивными) функциями нервной системы.
Методы исследования высших функции нервной системы
Многие из феноменов сознания трудны для экспериментального изучения, так как они, как полагают, присущи только человеку. В этом контексте напомним, что мозг макаки–резуса по массе составляет одну четвёртую часть мозга шимпанзе — ближайшего родственника человека по эволюционной линии, а масса мозга шимпанзе, в свою очередь, равна четверти массы мозга человека. Тем не менее, имеется много сходства между некоторыми нейрофизиологическими механизмами мозга обезьян и человека, и многие данные были получены в ходе лабораторных исследований приматов. Один из методов изучения мозга человека — стимуляция различных участков коры во время нейрохирургических вмешательств или стимуляция через имплантированные электроды. Второй метод — коррелирование клинических наблюдений деятельности мозга с данными аутопсии или, что, несомненно лучше, с наблюдениями над живым мозгом посредством позитронной эмиссионной томографии или ядерно-магнитной резонансной томографии. Применение этих и некоторых других методов позволяет изучать живой мозг не только во время чтения или слушания, но и в процессе активной умственной деятельности.
Некоторые формы обучения и памяти не являются уникальными для приматов или даже для позвоночных и, вероятнее всего, наблюдаются во всех частях нервной системы и у всех животных, имеющих нервную систему. Изучение молекулярных и клеточных проявлений некоторых феноменов (например привыкание, сенситизация и классический условный рефлекс) на нервной системе разных животных оказалось плодотворными и послужили началом развития молекулярной биологии обучения и памяти.
Обучение и память
Обучение— способность изменять поведение на основе опыта.Память— способность вызывать воспоминания о прошедших явлениях на сознательном или бессознательном уровнях. Оба процесса тесно взаимосвязаны.
Формы обучения
Различают ассоциативное и неассоциативное обучение. При неассоциативном обучении (привыкание и сенситизация) организм узнаёт только об одном стимуле, при ассоциативном (условный рефлекс) — об отношении одного стимула к другим.
Привыкание— наиболее простая форма обучения, во время которой нейтральный стимул повторяется много раз. Распространённой формой привыкания являетсяориентировочныйрефлекс(рефлекс «что такое?»). Ориентировочный рефлекс характеризуется реакцией внимания и генерализованным изменением электрической активности мозга. По мере повторения нейтральный стимул вызывает всё меньший и меньший электрический ответ. Наконец, субъект полностью привыкает к стимулу и игнорирует его (угасаниеориентировочногорефлекса).
Импринтинг(запечатлевание, вариант привыкания) — закрепление в памяти в раннем периоде развития отличительных признаков, воздействовавших на организм внешних объектов или некоторых врождённых поведенческих актов. Сохраняется в последующем и запечатлевает не индивидуальные, а видовые специфические характеристики жизненно важного объекта. Другими словами, животные обладают генетически закреплёнными сложными актами поведения, которые включаются после воздействия соответствующего раздражителя.
Сенситизацияявляется реакцией, противоположной привыканию. Повторные стимулы увеличивают ответ, если он сочетается один или несколько раз с приятным или неприятным раздражителем. Общеизвестно увеличение интенсивности жизненно важных пробуждающих стимулов у людей (спящая мать в шумной обстановке мгновенно просыпается, услышав голос своего ребенка).
Условныйрефлекс(это понятие в научный оборот ввёлИван Петрович Павловв 1901 г.) — рефлекторный ответ на индифферентный (т.е. не вызывавший ранее реакции организма) стимул, который, будучи несколько раз повторён вместе с другим стимулом, всегда вызывающим реакцию организма, сам вызывает эту реакцию. В классических опытах И.П. Павлова слюноотделение у голодной собаки вызывали кормлением собаки мясом. Затем перед каждым кормлением звенел звонок, и эту процедуру повторяли до тех пор, пока только звонок (без последующего кормления мясом) стал вызывать слюноотделение. В этом эксперименте мясо —безусловныйраздражитель(всегда вызывает слюноотделение), а звонок —условныйраздражитель.
В медицинской практике огромное количество соматических, висцеральных и другого рода патологических изменений может возникнуть именно на условно–рефлекторной основе. Сочетание висцеральных ответов с условными стимулами — биологическаяобратнаясвязь. Попытки использовать условно-рефлекторные воздействия для восстановления нарушенного сердечного ритма или для снижения АД не смогли стать альтернативой традиционной терапии из-за слишком малых величин депрессорных ответов, получаемых этим способом, но этот приём используется в ряде областей практической медицины.
Исчезновениеусловногорефлекса. Если условный раздражитель повторяется без подкрепления безусловным стимулом, то условный рефлекс, в конце концов, исчезает. Этоугасание,иливнутреннееторможение. Если немедленно после условно–рефлекторного раздражения появляется какое–либо неожиданное внешнее воздействие, то условный рефлекс не возникает. Это —внешнееторможение. Однако если условный рефлекс время от времени поддерживается сочетанием условного стимула со стимулом безусловным, то он сохраняется длительное время. Важно, чтобы безусловный стимул был ассоциирован с активацией системы поощрения (или системы наказания и боли), чтобы стимулировать положительное или отрицательное подкрепление условного рефлекса.
Инструментальныерефлексы. Самостоятельной формой условных рефлексов являются инструментальные рефлексы. Они формируются на основе активной и целенаправленной деятельности. Сюда же относится дрессировка, илиоперантноеобучение(обучение методом «проб и ошибок»).
Физиологическоезначениеусловныхрефлексов. Условные рефлексы:вырабатываются и накапливаются в индивидуальной жизни каждого субъекта,носят приспособительный характер, делая поведение наиболее пластичным, приспособленным к конкретным условиям среды;имеют сигнальный характер, т.е. предшествуют, предупреждают последующее возникновение безусловно рефлекторных реакций, подготавливая к ним организм. Таким образом, условные рефлексы — индивидуально приобретённые системные приспособительные реакции, возникающие на основе образования в нервной системе временной связи между условным (сигнальным) раздражителем и безусловно рефлекторной деятельностью.
Участиеобоихполушарийвмеханизмеобразованияусловногорефлекса. Выработка условного рефлекса у обезьян и кошек в ответ на зрительный стимул с одним закрытым глазом и тестирование образовавшегося условного рефлекса на другом (ранее закрытом) глазу показало, что ранее закрытый глаз воспроизводит условный рефлекс. Условный рефлекс образуется и воспроизводится в условиях перерезанного зрительного перекреста, когда зрительная информация от каждого глаза поступает только в ипсилатеральные отделы коры. Если вдобавок к нарушению зрительного перекреста пересечены передние и задние комиссуры и мозолистое тело, то переноса условно–рефлекторной реакции не наблюдается. Подобные результаты были получены и на людях с врождённым отсутствием мозолистого тела и на пациентах, у которых полностью удалялось мозолистое тело с целью облегчения тяжёлых эпилептических приступов. Эти данные демонстрируют, что обучение, осуществляемое при наличии одного глаза, передаётся механизмом нервного кодирования другому глазу через комиссуры полушарий. Имеются доказательства, что подобного рода передача информации осуществляется и через другие сенсорные пути. Так, наблюдения над больными, у которых не полностью перерезано мозолистое тело, показали, что зрительная информация передаётся через дорсальную часть мозолистого тела, в то время как передние отделы имеют отношение к передаче слуховой и соматосенсорной информации.
Память
Существует несколько форм памяти: от примитивных (типа привыкания и сенситизации) до комплекса сознательных воспоминаний. Формы памяти (в зависимости от типа сохраняемой информации) подразделяют на недекларативную (рефлексивную, безотчётную) и декларативную (табл. 17–1).
Таблица 17–1. Формы памяти
|
Недекларативнаяпамять(рефлексивная, безотчётная) |
|
Привыкание Сенситизация Классический условный рефлекс Мастерство (навыки) и привычки Запечатлевание (импринтинг) |
|
Декларативнаяпамять(объяснимая, узнаваемая) |
|
Явления, события, факты, полученный опыт Объяснение жизненного опыта, самоанализ и контроль |
Недекларативная память в большинстве случаев полностью бессознательна. Декларативная память всегда вовлекает сознание для воспроизведения событий и фактов. Эти виды памяти образуются различными путями и в различных частях мозга. Вне зависимости от форм памяти, понимание механизмов её фиксации и сохранения не выходит за рамки гипотез.
Декларативная память, связанная с моторной активностью, может превращаться в рефлексивную память посредством постоянного повторения. Например, гимнасты или цирковые акробаты так тренируют движения, что их ответы в результате тренировки становятся практически «инстинктивными». Подобным же образом многие аспекты поведения в комплексе вождения автомобиля или игры на фортепиано приобретают черты привычных ответов.
Декларативную память принято классифицировать как мгновенную (сенсорную), кратковременную, промежуточную долговременную и долговременную.
Мгновеннаяпамять— следовый отпечаток действующего стимула в рецептирующей структуре. Она продолжается доли секунды, не зависит от воли и не может быть подвергнута сознательному контролю.
Кратковременнаяпамятьпродолжается от нескольких секунд до нескольких минут. Обычно это соответствует семи–десятизначному номеру телефона или от 7 до 10 отдельных коротких фактов. Кратковременная память продолжается только то время, пока человек продолжает думать о телефонном номере или полученных фактах. Предполагают, что в основе кратковременной памяти находитсяповторнаямногократнаяциркуляциянервныхимпульсовпозамкнутойсистеменейронов.
Другое возможное объяснение кратковременной памяти — пресинаптическоеоблегчение. Нейромедиаторы, секретируемые терминалями, вызывают продолжительное облегчение (в зависимости от вида нейромедиатора) — от нескольких секунд до нескольких минут. Нейронные круги этого типа могут приводить к кратковременной памяти.
Ещё одно гипотетическое объяснение кратковременной памяти — синаптическаяпотенциация, которая может увеличивать синаптическое проведение в результате накопления больших количеств ионов кальция в пресинаптических терминалях. Когда количество ионов кальция становится больше возможности их абсорбции митохондриями и эндоплазматической сетью, то избыток ионов кальция приводит к продолжительному пресинаптическому выделению нейромедиатора.
Промежуточнаядолговременнаяпамятьможет сохраняться от нескольких минут до нескольких недель. Если эта память случайно не утрачивается, и её следы становятся более постоянными, то она переходит в разряд долговременной памяти. Полагают, что сохранение этой памяти —результатвызванныхактивациейметаботропныхрецептороввременныхфизико-химическихизмененийвпостсинаптическихнейронах.
Эксперименты на морских моллюсках Aplysiaпоказали, что промежуточная долговременная память является результатом временных физико–химических изменений в пресинаптических терминалях и постсинаптической мембране. На рис. 17–0 показана схема этих экспериментов.
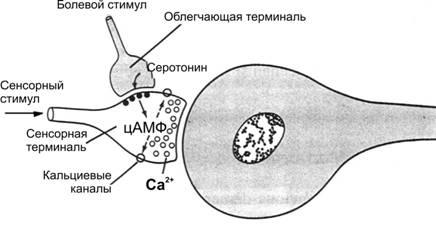
Рис. 17–0. Система памяти у улитки Aplysia.
На рисунке представлены две пресинаптические терминали. Одна терминаль от сенсорного нейрона оканчивается на поверхности стимулируемого нейрона — сенсорнаятерминаль. Другая терминаль, являющаяся пресинаптическим окончанием, лежит на поверхности сенсорной терминали, этооблегчающаятерминаль. Повторная стимуляция сенсорной терминали вызывает постепенное уменьшение потенциала нейрона вплоть до полного его исчезновения. Возникшеепривыканиепоявляется из-за малой биологической значимости сигналов. Напротив, если болевые стимулы раздражают облегчающую терминаль каждый раз, когда стимулируется сенсорная терминаль, то сигналы в постсинаптическом нейроне становятся всё более и более сильными и продолжают оставаться сильными в течение минут, часов, дней и даже 3 нед без дальнейшей стимуляции облегчающей терминали. Возможный молекулярный механизм промежуточной долговременной памяти заключается в следующем: облегчающая терминаль выделяетсеротонинк мембране сенсорной терминали, серотонин взаимодействует с серотониновыми рецепторами сенсорной терминали, и эти рецепторы активируют аденилатциклазу внутри мембраны, приводя к образованию цАМФ. цАМФ активирует протеинкиназу, которая через цепь реакций инактивирует калиевые каналы в мембране сенсорной терминали и приводит к блоку калиевой проводимости мембраны нейрона (эта блокада может продолжаться от нескольких минут до нескольких недель). Нарушение K+–проводимости увеличивает продолжительность ПД. Продолжительный ПД интенсивно активирует кальциевые каналы, вызывая накопление кальция внутри сенсорной терминали, Ионы кальция увеличивают выделение медиатора, вызывая тем самым облегчение синаптической передачи. Таким образом,ассоциативныйэффектстимуляцииоблегчающегонейронаодновременносостимуляциейсенсорногонейронавызывает продолжительное возбуждение сенсорной терминали и устанавливает следы памяти.
Долговременнаяпамятьзависит, как полагают, от реструктуризации синапсов, что приводит к долговременным изменениям их чувствительности при передаче нервных сигналов. По настоящее время мы не знаем, что именно является той матрицей (нейронные сети, макромолекулы или их комплексы в нейронах), что фиксирует память и как происходит извлечение следов памяти.
Консолидацияпамяти. Для того чтобы кратковременная память превратилась в долговременную память и могла быть востребована через недели и годы, требуется её консолидация. Минимальный срок для консолидации — 5–10 мин, для прочной консолидации — 1 час и более. Так, если мозгу предъявлено сильное сенсорное воздействие, а вслед за ним (через единицы минут) следует электрошок, то сенсорное воздействие не запоминается. Такой же эффект оказывают общая анестезия, контузия мозга и другие воздействия. Однако если электрошок применяется через 15–20 мин, то часть памяти в последующем может быть воспроизведена. Через один час память почти полностью консолидируется, и электрошок не нарушает её.
Повторениеинформации. Для консолидации памяти важно повторное предъявление одной и той же информации. Этим можно объяснить намного лучшее вспоминание небольшого количества глубоко изученного материала, чем большого количества поверхностно усвоенной информации (классический пример — студент перед экзаменом). Бодрый и ясно мыслящий человек может консолидировать память намного лучше, чем человек, находящийся в состоянии умственной усталости.
Кодированиепамяти. Консолидация памяти сопровождается кодированием новой памяти на различные классы. Во время консолидации новая память не складывается в порядке поступления в мозг, а поступает на хранение в прямой ассоциации с формами памяти того же самого вида. При этом предварительно происходит сравнение старой и новой памяти на предмет выяснения сходств и различий. Это обстоятельство позволяет находить необходимую информацию, используя последние сведения для извлечения более ранних данных. Кодированию памяти предшествуют процессы рабочей памяти.
Рабочаяпамять— временный запас информации, используемый для планирования будущих действий — прямо не связана (в отличие от консолидации памяти) с гиппокампом, а является функцией разных областей новой коры и в первую очередь — префронтальной области (см. ниже). Когда испытуемый говорит, вспоминая пространственное расположение зрительных стимулов, происходит увеличение активности в правой затылочной, теменной и лобной долях коры. Попытка испытуемого вспомнить последовательность букв алфавита и повторять их «в уме» (не называя), увеличивает активность в других областях коры. Префронтальная область разделена на отдельные сегменты для хранения различных видов временной памяти (например, хранилища для временной памяти на размеры и форму объектов, на размеры частей тела или его движения).
Анатомическаятопографияпамяти
Гиппокамп. В процессы кодирования при превращении кратковременной памяти в память долговременную вовлекаются гиппокамп и прилежащие к нему части медиальной височной коры (рис. 17–1А).
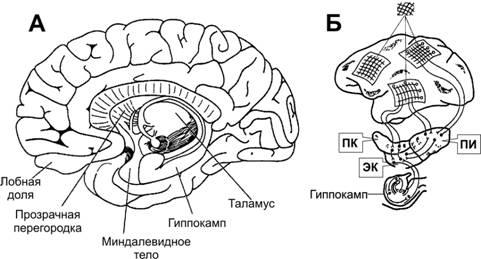
Рис.17–1.ДЕКЛАРАТИВНАЯПАМЯТЬ.А.Областимозга,имеющиеотношениеккодированиюдолговременнойпамяти.Б.Путикодированиядекларативнойпамятивмедиальнойвисочнойобласти. Вход информации из зрительной и других областей коры через парагиппокампальную извилину (ПИ), периринальную кору (ПК) и энторинальную кору (ЭК) в гиппокамп и возвращение кодированной информации в кору (помечены заштрихованными областями.
Билатеральное разрушение вентрального гиппокампа или болезненный процесс, поражающий нейроны гиппокампа, вызывают поразительный дефект в памяти на последние события. С таким поражением гиппокампа человек сохраняет интактными промежуточную и долговременную память, как и недекларативную (безотчётную) память. Люди с таким поражением адекватно с позиций сознательной памяти осуществляют попытки показать, как они концентрируются на том, что они собираются делать, но если они отвлекаются даже на очень короткий период времени, вся память о том, что они делали или что они собирались делать, исчезает. Они могут обучаться новому материалу и помнить о том, что было до заболевания, но они не способны формировать новую долговременную память.
Сосцевидныетелаиталамус, прямо и опосредованно связанные с гиппокампом, также вовлечены в механизмы кратковременной памяти.
Так, люди с алкогольным поражением мозга теряют память на последние события, что коррелирует с патологическими изменениями в сосцевидных телах, имеющих обширные эфферентные связи с гиппокампом. Сосцевидные тела связаны и с таламусом, поэтому поражение таламуса также вызывает потерю кратковременной памяти. Из таламуса волокна, имеющие отношение к памяти, проецируются к лобной коре и оттуда к основанию переднего мозга. От основания переднего мозга идут диффузные холинергические проекции ко всем отделам новой коры, миндалевидному телу и гиппокампу. Миндалевидное тело тесно контактирует с гиппокампом, но прямого отношения к кодированию декларативной памяти не имеет. Активность миндалевидного тела добавляет к воспоминаниям эмоциональную окраску.
Корабольшихполушарий. В то время как кратковременная память кодируется гиппокампом и функционально связанными с ним структурами, хранение долговременной памяти происходит в различных отделах новой коры.
Стимуляция разных участков новой коры во время нейрохирургических операций вызывает детальные воспоминания о событиях далёкого прошлого. Очевидно, различные элементы воспоминаний — зрительные, обонятельные, слуховые и др. — располагаются в регионах коры, соответствующих этим функциям, и каким-то образом отдельные фрагменты памяти собираются вместе, когда они извлекаются из хранилищ памяти. Стимуляция некоторых участков височной доли коры вызывает изменения в оценке окружающих людей. Например, при применении стимула субъект мог ощущать себя чужим в семье или мог чувствовать, что сейчас произойдёт что-то такое, что уже случалось ранее. Появление чувства бесцеремонности или отчуждённости в соответствующих ситуациях, вероятно, помогают нормальному человеку приспосабливаться к окружению. Среди чужих людей он насторожен и бдителен, в то время как в семейной обстановке бдительность ослабляется. Неуместное чувство преувеличенной бесцеремонности к новым событиям или окружающим клинически известно как «dejаvu— уже виденное». Этот феномен возникает время от времени и у нормальных индивидуумов, а также может наблюдаться как аура (ощущение, предшествующее наступлению судорожного припадка) у больных с височно-лобной эпилепсией.
Возможныймеханизмкодированиядекларативнойпамятив форме рабочей гипотезы представлен на рис. 17–1Б. Согласно гипотезе, входящие сенсорные стимулы, инициируют в адекватных сенсорному сигналу ассоциативных областях коры больших полушарий возбуждённое состояние нейронных цепей. Нервные сигналы направляются к медиальной височной доле, к парагиппокампальной извилине, периринальной и энторинальной областям. Из энторинальной коры сигналы поступают в гиппокамп, где происходят пока ещё неизвестные механизмы кодирования кратковременной памяти. Пути, выходящие из гиппокампа через энторинальную кору соединяются вместе с нейронными цепями новой коры (откуда поступили сигналы) и образуют постоянную и стабильную долговременную память, которая может быть извлечена различными пусковыми сигналами.
НарушенияпамятиприболезниАльцхаймера. Основной признак болезни Альцхаймера — исчезновение нейронов в тех частях лимбической системы, которые обеспечивают процессы переноса памяти. Болезнь Альцхаймера характеризуется прогрессивной утратой памяти и функций познания. Ранние проявления болезни Альцхаймера состоят в уменьшении метаболизма и кровотока в теменной коре с обеих сторон. Одновременно утрачиваются холинергические волокна коры больших полушарий, базальных и релейных ядрах, посылающих волокна к гиппокампу и миндалевидному телу. Причина дегенерации холинергических нейронов неизвестна. Цитологически внутри нейронов обнаружено нарушение образования белков, связанных с микротрубочками — формируются сенильныебляшки, содержащиеb–амилоид, нарушающий структуру нервных волокон и глиальных клеток. Таким образом, болезнь Альцхаймера — метаболическое дегенеративное заболевание.