Экология организмов Вопросы к 1 коллоквиуму Обзор основных групп микроорганизмов и особенности их экологии
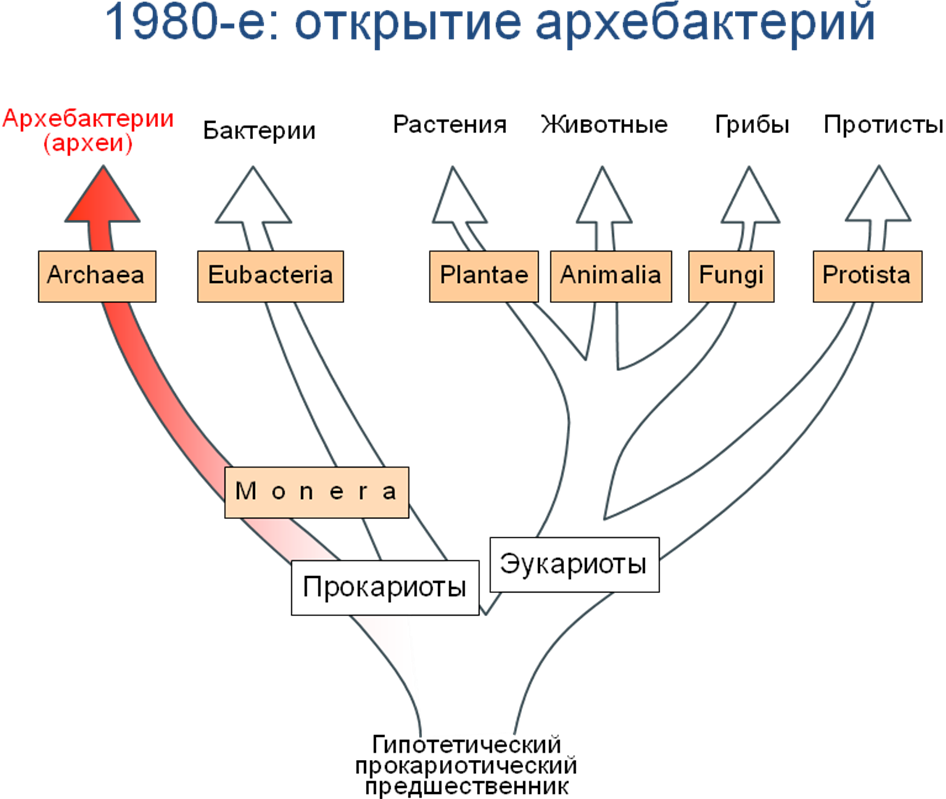
Изменение взглядов на разнообразие организмов в последние десятилетия: переход от двуцарственной системы к многоцарственной
Под термином мегасистематика понимается классификация живых организмов на самом высоком таксономическом уровне, то есть уровне царств (надцарств, подцарств) и выделяемых в последнее время некоторых более высоких таксономических категорий, таких как доминионы и империи. Впервые столь глобальное подразделение живых существ было предпринято Аристотелем (ок. 384-322 до н.э.), который предложил различать растения и животные. Критериями для отнесения живых организмов к той или другой группе служили, согласно Аристотелю, подвижность и чувствительность. Растения неподвижны и нечувствительны, животные - наоборот. Надо отметить, что уже сам Аристотель столкнулся с трудностями при классификации некоторых прикрепленных или малоподвижных организмов, таких как губки, голотурии и асцидии. Он полагал, что в них смешана животная и растительная природа. Несмотря на эти затруднения, классификация Аристотеля была принята Карлом Линнеем (1707-1778), который в своей знаменитой "Systema Naturae" (1735), выделил два царства: Vegetabilia (растения) и Animalia (животные). Впоследствии к двум критериям различения растительных и животных организмов Аристотеля Жан Батист Ламарк (1744-1829) добавил еще и способ питания - автотрофный для растений и гетеротрофный для животных. Такая двуцарственная система живого просуществовала практически до наших дней, хотя время от времени и подвергалась сомнению. Осложнения стали накапливаться еще со времени открытия Левенгуком (1632-1723) мира микроскопических организмов, названных им анималькулами. Само название указывало на причисление этих живых существ к царству животных, что основывалось на критерии подвижности. К.Линней в своей системе все известные к тому времени микроскопические организмы объединил в рамках рода Chaos, который включался в отряд Zoophyta, наряду с медузами, головоногими моллюсками, иглокожими и др. Само название Zoophyta указывает на двойственность, по мнению Линнея, природы всех этих живых существ, которых знаменитый шведский натуралист все же относит к царству животных. По мере развития микроскопической техники выяснилось, что среди анималькул есть формы, снабженные хлоропластами, то есть растительные по своей природе, есть питающиеся гетеротрофно, принадлежащие царству животных, и, наконец, имеются отличные от тех и других микроорганизмы (монеры). Всех их Геккель (Haeckel, 1866) предложил выделить в самостоятельное царство Protista (протисты), к которому он также отнес бактерий и некоторых низших многоклеточных животных (губки). Эта точка зрения не нашла поддержки, и в более поздних работах сам Геккель отказался от нее, распределив протистов между царствами растений и животных. Однако противоречивость двуцарственного подразделения живого становилась все более и более очевидной. Свое выражение это находило в практической систематике, в рамках которой ботаники и зоологи относили (и до сих пор относят!) одни и те же организмы к разным таксонам. К примеру, согласно ботанической номенклатуре эвгленовые выделяются в самостоятельный отдел (тип), динофитовые (динофлагелляты) в подотдел или, по крайней мере, в класс, а вольвоксовые в класс или порядок. Зоологи же рассматривают всех их лишь как отряды в классе жгутиконосцев. Таких "двуцарственных" видов, которые классифицируются одновременно как в качестве растений, так и в качестве животных, насчитывается сейчас около 30 тысяч (Кусакин, Дроздов, 1998). Более того, во многих группах водорослей (криптофитовые, динофитовые, золотистые, зеленые) имеются бесцветные, то есть лишенные хлоропластов формы. Их, следуя критерию способа питания, следовало бы относить к животному царству, хотя их родство с остальными представителями соответствующих таксонов, обладающих хлоропластами, несомненно и никогда ни у кого сомнения не вызывало. Можно вспомнить в этой связи и классический пример некоторых эвгленовых, которые на свету питаются автотрофно, а в темноте переходят к гетеротрофному способу питания. Несмотря на парадоксальность ситуации, подобная система классификации с молчаливого согласия ботаников и зоологов существовала вплоть до самого недавнего времени, да и поныне еще принята во многих учебных курсах, руководствах и пособиях. Одна из причин подобного положения дел - мелкие размеры простейших живых организмов, что до появления электронного микроскопа резко ограничивало набор признаков, используемых при таксономических построениях. Ситуация стала постепенно изменяться начиная с 60-х годов, когда в связи с активным внедрением в биологию методов электронной микроскопии (особенно интенсивно эти исследования проводились в 70-80-х годах) стали накапливаться принципиально новые данные по тонкому строению (ультраструктуре) простейших живых организмов. Оказалось, что на этом уровне выявляются достаточно отчетливые морфологические признаки (тонкое строение покровов, жгутикового аппарата, митохондрий, хлоропластов и др.), которые можно использовать в качестве надежных критериев при выяснении степени родства организмов. Другая волна новой информации стала быстро распространяться с 80-х годов со стороны молекулярной биологии, когда появилась возможность сравнивать степень сходства нуклеиновых кислот разных организмов. При выполнении такого рода исследований при помощи специальных методик проводится так называемый сиквенс, то есть определяется последовательность нуклеотидов в том или ином участке РНК или ДНК. Затем сиквенсы одних и тех же участков нуклеиновых кислот разных организмов сравниваются друг с другом. Число выявленных при этом различий определяет степень родства сравниваемых организмов: чем больше несовпадений в последовательностях нуклеотидов, тем дальше отстоят они друг от друга. Для таксономических построений используются преимущественно рибосомальные РНК (5S рРНК, 16-18S рРНК, 23-28S рРНК), в ряде случаев митохондриальная ДНК и последовательности ядерной ДНК, кодирующие определенные белки (а- и b- глобулины, стрессовые или хит-шоковые белки, цитохромы и др.). Сразу следует оговориться, что считать молекулярно-биологические данные истиной в последней инстанции и панацеей от всех бед, что нередко практикуется, нельзя ни в коем случае. Все эти построения зиждутся на признании, в той или иной форме, гипотезы о нейтральности эволюции и следующего из нее принципа молекулярных часов (Кимура, 1985). Согласно последнему, замены нуклеотидов (то есть точечные мутации) в ходе эволюции происходят с постоянной частотой, одинаковой для разных групп организмов. Это-то и дает основание для определения степени родства между ними при сравнении сиквенсов гомологичных участков нуклеиновых кислот. Более того, молекулярные часы можно калибровать. Для этого достаточно вычислить число замен нуклеотидов в определенных участках нуклеиновых кислот двух организмов (или большего их числа), время дивергенции которых известно из палеонтологической летописи. Затем результаты такой калибровки можно переносить на любые другие группы организмов. В действительности скорость эволюции, а соответственно и скорость накопления мутаций, не остается постоянной даже в пределах одной филогенетической линии (Татаринков, 1987; Тыщенко, 1992). Периоды интенсивного протекания эволюционных изменений сменяют, как правило, длительные периоды застоя (стазиса). Эти представления учитываются в современных молекулярных работах. Для анализа выбирают, как правило, более-менее консервативные типы или участки нуклеиновых кислот, параллельно используются результаты сопоставления сиквенсов разных типов или участков ДНК и/или РНК. Несмотря на это, к молекулярно-биологическим данным следует относится с известной осторожностью и при их интерпретации учитывать традиционные морфологические признаки. Так или иначе, но именно ультраструктурные и молекулярно-биологические данные привели к той революции в мегасистематике живого, которую мы переживаем в настоящее время. Во-первых, было выявлено несомненное родство между группами, родственные связи между которыми ранее даже не предугадывались. Так. например, среди водорослей эвгленовые оказались близкородственны группе паразитических жгутиконосцев-кинетопластид (к ним, в том числе, относятся трипанозомы - возбудители таких опасных заболеваний, как сонная болезнь, болезнь Чагоса и др.), а динофитовые - инфузориям и споровикам, всегда считавшимися зоологическими объектами и не обнаруживавшими на светооптическом уоовне ни малейшего сходства как с динофитовыми, так и между собой Во-вторых, различия между представителями пазных групп протистов оказались, по крайней мере, не меньше, а зачастую и больше, чем между типичными растениями и животными. Наметился кризис в классических представлениях о мега-системе живого (Кусакин, 1995), который до определенной степени напоминает создавшийся в физике 20-х годов вслед за открытием новых свойств электрона. Путь к его разрешению видится в создании многоцарственных систем живых организмов, различные варианты которых можно найти в сводках последних десятилетий (Whittaker, 1969; Cavalier-Smith, 1981, 1999; Corlis, 1987; Старобогатов, 1986; Серавин, 1989; Карпов, 199о! 2000; Кусакин, 1995; Кусакин, Дроздов, 1998 и др.). Однако, прежде чем приступить к их анализу, следует обсудить некоторые общие принципы, на которых может базироваться современная мегасистема живого.
Особенности развития экологии во второй половине XX в.
Математическое моделирование
Компьютеризация. Базы данных. Количественные оценки. Прогнозы
Международное сотрудничество
Прикладная экология. Охрана природы
Организационные мероприятия (общества, журналы, совещания)
Конец XX века
Катастрофический рост численности населения
Технологический взрыв. Приоритет технического развития в ущерб гармонии с природой и совершенствованию культуры.
Иллюзия независимости от природы и все большая зависимость не деле (ресурсы).
Окончательное осознание ограниченности ресурсов (не только учеными, но и политиками)
Проблема охраны природы
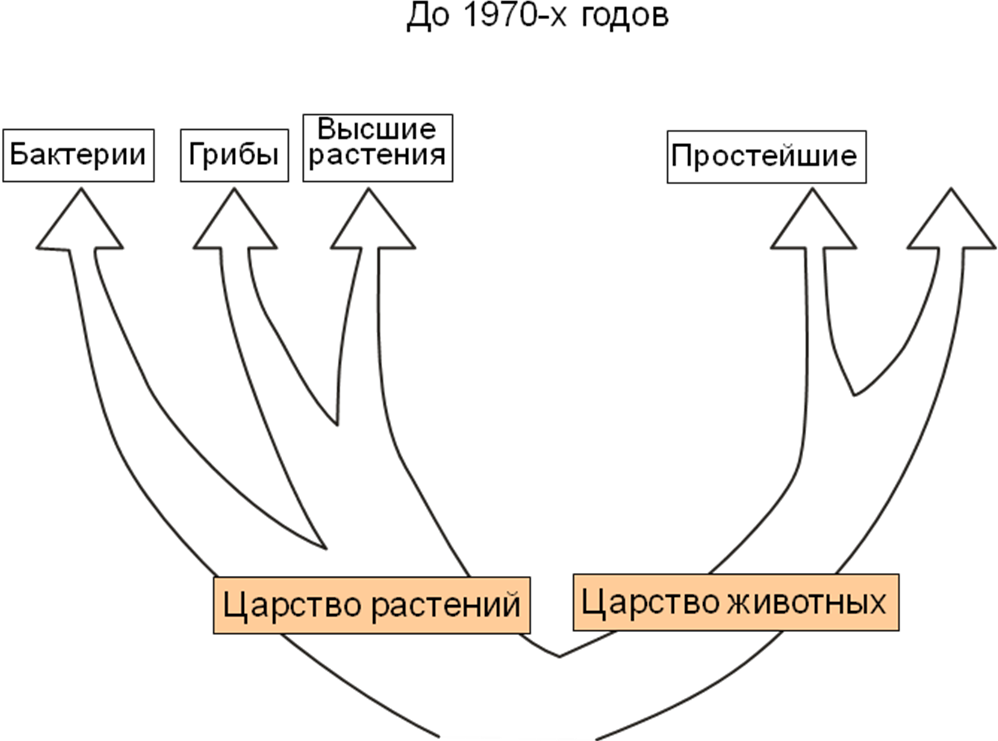
Двухцарственная система:
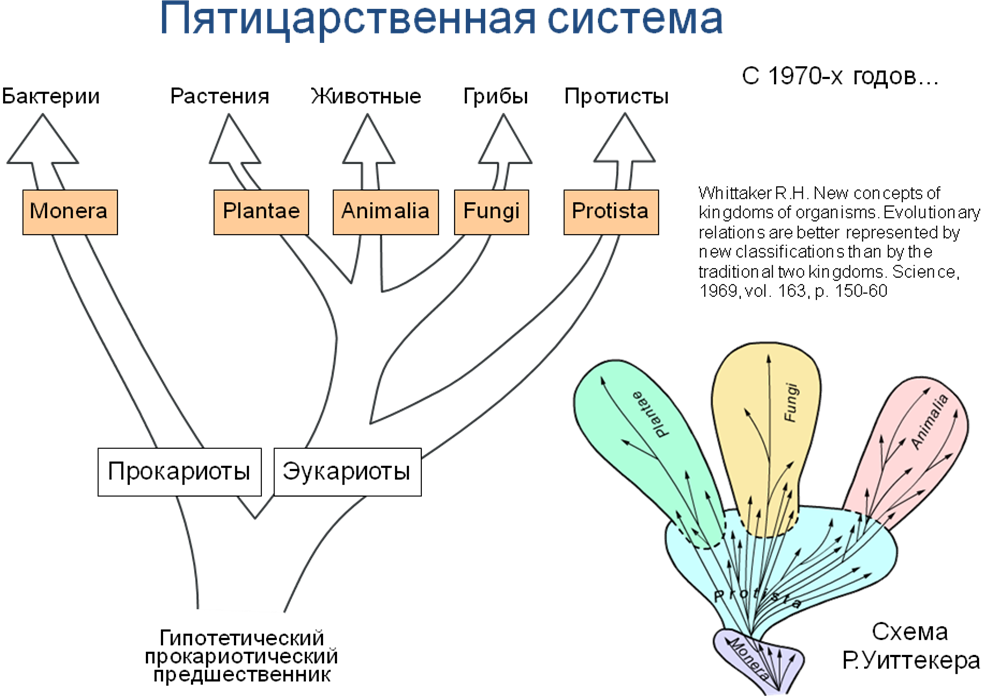 Пятицарственная
Пятицарственная