

11.1.2. Общие свойства мышечных тканей
|
Несмотря на различие строения мышечных тканей, можно указать следующие общие моменты. |
|
Принцип сокращения |
а) Во всех этих тканях в процессе сокращения происходит
б) За счёт этого уменьшается длина волокна или клетки. |
|
Участие Са2+ |
Для протекания вышеуказанного процесса необходимо
что происходит в ответ на нервное воздействие. |
|
Энерго- обеспече- ние |
а) Для энергетического обеспечения сокращения мышечные клетки или волокна, как правило, содержат б) Кроме того, они в большей или меньшей степени способны создавать
|
|
АТФ |
а) Непосредственным источником энергии при сокращении миофибрилл является АТФ (аденозинтрифосфат). б) АТФ
в) При этом конкретными структурами, которые
являются т.н. головки белка миозина (см. ниже), составляющего толстые миофиламенты. |
|
Креатин- фосфат |
В скелетной и сердечной мышечных тканях, помимо АТФ, функцию аккумулятора энергии может выполнять ещё одно вещество – креатинфосфат. Оно
|
|
Базальная мембрана |
а) Наконец, отметим. что
покрыты базальной мембраной. б) В связи с этим следует упомянуть термин "сарколемма"; под ним понимают:
|
|
Теперь более подробно рассмотрим каждую мышечную ткань – скелетную, сердечную и гладкую. |
![]()
11.2. Скелетная поперечнополосатая мышечная ткань
![]()
11.2.1. Мышечные волокна на светооптическом уровне
|
а) Итак, скелетная мышечная ткань состоит из мышечных волокон, которые
б) Используя в качестве иллюстрации снимки с препаратов языка, перечислим ещё ряд характерных свойств мышечных волокон. |
![]()
11.2.1.1. Основные светооптические характеристики
I. Размеры и тинкториальные свойства волокон
|
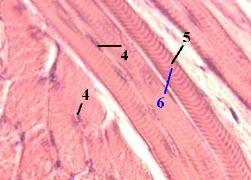
1,а-б. Препарат - поперечнополосатая скелетная мышечная ткань; срез языка. Окраска гематоксилин-эозином. | ||
|
Ориента- ция мышечных волокон |
а) Предварительно заметим, что в языке группы волокон идут в нескольких (почти взаимно перпендикулярных) направлениях. б) Поэтому на препаратах одни мышечные волокна срезаны продольно (1), а другие – поперечно (2). |
а) Малое увеличение
Полный размер |
|
1. Размеры волокон |
а) Обычный диаметр мышечных волокон – 50-70 мкм, что почти в 10 раз больше диаметра эритроцита (7,5 мкм; п. 8.2.1.1). б) Длина же волокна (а одновременно и миосимпласта) совпадает с длиной соответствующей мышцы, т.е.
| |
|
2. Тинкто- риальные свойства |
а) Мышечные волокна отличаются
они интенсивно красятся эозином в ярко-розовый цвет. б) Причина оксифилии – высокое содержание белков. в) Для сравнения напомним, что оксифильными являются и пучки коллагеновых волокон в плотных волокнистых соединительных тканях; тема 9). | |

II. Ядра мышечных волокон
|
3. Ядра мышеч- ного волокна |
а) Ядра удобней изучать на продольных срезах мышечных волокон. б) При этом
|
б) Среднее увеличение
Полный размер |
|
в) Ядра симпласта (4) имеют узкую, палочковидную форму, ядра же миосателлитов - овальные, но их трудно различить среди ядер симпласта. г) Количество ядер в каждом миосимпласте очень велико,
| ||
|
4. Центриоли |
а) В миосимпластах отсутствуют центриоли. б) Поэтому ядра не способны делиться – ни в нормальном состоянии мышечного волокна, ни при регенерации. | |
III. Миофибриллы и поперечная исчерченность
|
5. Мио- фибриллы |
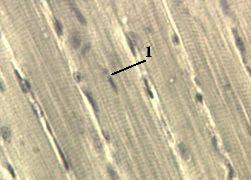
а) Как уже отмечалось, миофибриллы (7) занимают около 70% объёма миосимпласта. б) Различить их можно в поперечно срезанных мышечных волокнах при большом увеличении светового микроскопа. |
в) Большое
увеличение |
|
в) Они имеют вид точек, которые заполняют почти всё сечение миосимпласта. г) Диаметр миофибриллы – 1,5 мкм. д) На поперечном сечении мышечного волокна содержится около 1400 миофибрилл. | ||
|
6. Попереч- ная исчерчен- ность |
а) Сами миофибриллы на светооптическом уровне видеть ещё можно, но различить в них поперечную исчерченность на этом уровне уже нельзя. б) Однако можно наблюдать поперечную исчерченность мышечных волокон (или, точнее, миосимпластов). | |
|
в) Так, на снимке (1,б) у продольно срезанных волокон заметно регулярное чередование |
| |
|
г) Ещё лучше выявляется данная исчерченность (и другие перечисленные выше особенности) при окраске препарата железным гематоксилином. | ||
 Полный
размер
Полный
размер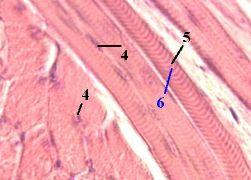
IV. Окраска железным гематоксилином
|
2. Препарат - поперечнополосатая скелетная мышечная ткань, срез языка. Окраска железным гематоксилином. | |
|
У продольно срезанных волокон хорошо видны три основные особенности скелетной мышечной ткани, а именно то, что
|
Полный размер |
![]()