

Лидерная м-рнк uuuuuuuu- он 3′

Рис.13. Последовательность оснований на аттенюаторе trр-оперона.


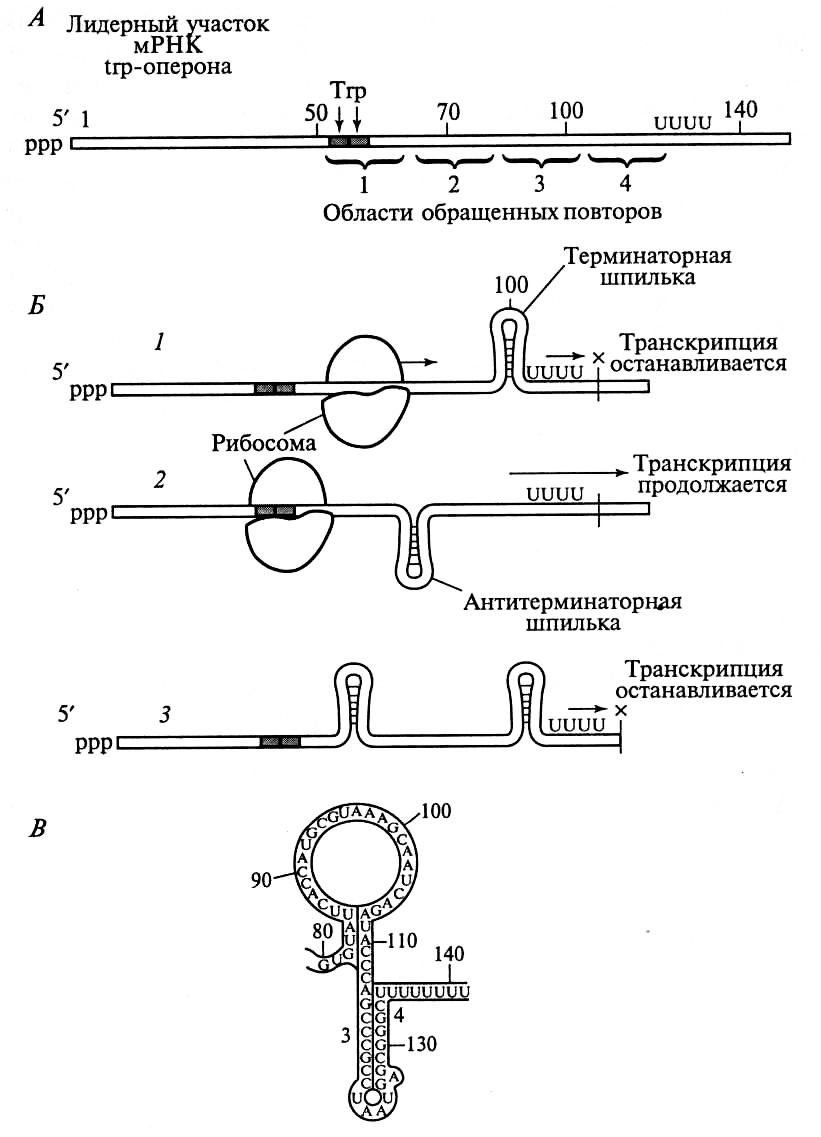

Рис.14. Процесс регуляции
транскрипции с участием
рибосом: А-лидерный
участок мРНК, Б-схема
регуляции; В-«шпилька»
на мРНК.
Образование терминаторной шпильки вызывает остановку транск- рипции на участке, содержащем подряд несколько остатков урацила. Остановка транскрипции на участке аттенюаторе называется аттенуацией. РНК-полимераза отсоединяется от ДНК и гены не считывается.

Рис.15. Образование «шпильки» на 3′-конце м-РНК.
Таким образом, большая концентрация триптофана в клетке, позволяет этой аминокислоте быстро включаться в лидерный пептид и, обеспечив быстрое и плавное движение рибосомы по мРНК, блокировать транскрипцию генов, контролирующих ее собственный синтез через аттенюаторный механизм.
В условиях низкой концентрации триптофана или его отсутствии (триптофановое голодание) при синтезе лидерного пептида рибосома задерживается на двух рядом расположенных кодонах УГГ (рис.16-17).


Met – Lys – Ala – Ile – Phe – Val – Leu – Lys – Gly – Trp –Trp – Arg – Thr – Str – стоп
АУГ-ААА-ГЦА-АУУ-УУЦ-ГУА-ГУГ-ААА- ГГУ- УГГ- УГГ- ЦГЦ -АЦУ-УЦЦ-УГА
Рис.16. Последовательность аминокислот в лидерном пептиде trр и
последовательность оснований в мРНК
Застрявшая рибосома, отстает от РНК-полимеразы. Это отставание влияет на синтезируемую и-РНК так, что формируется антитерминаторная шпилька, которая препятствует образованию терминаторной шпильки и РНК-полимераза проскакивает аттенюатор, продолжая транскрипцию генов; активно синтезируются ферменты необходимые для пополнения запаса триптофана в клетке.
Итак, синтез триптофана может регулироваться trp-опероном через аттенюаторный механизм, который чутко реагирует на изменение концентрации этой аминокислоты в клетке.
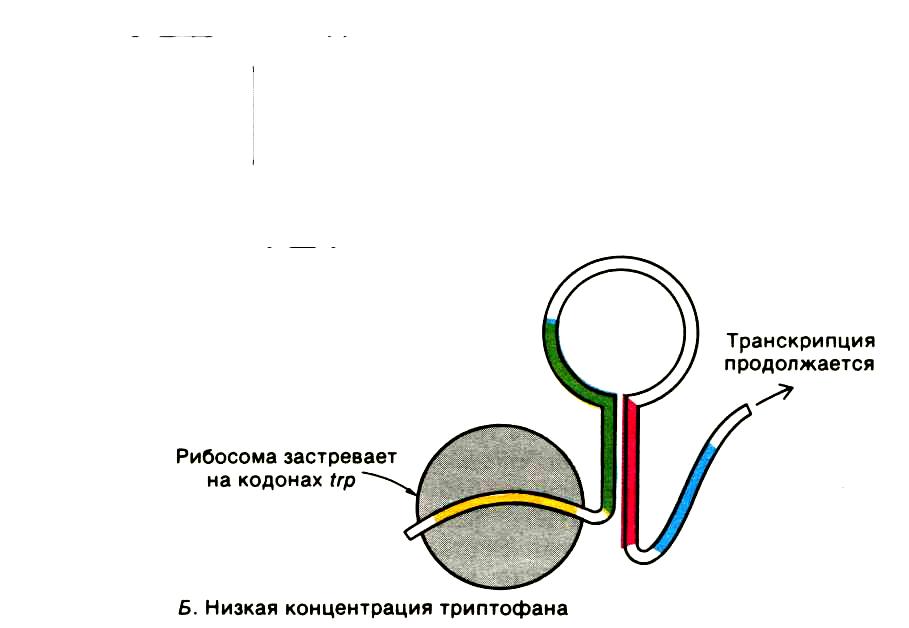
Рис.17. Схема аттенюации trр-оперона Е.соli при низкой
концентрации триптофана
1.5. Гены эукариотических организмов
Гены эукариотических организмов устроены сложнее, чем гены прокариот (рис.18). Регуляторная часть эукариотических генов состоит из промотора, терминатора, энхансера и сайленсера. Промоторы генов эукариот кодирующих белки, расположены левее точки начала транскрипции (+1) и включают нескольких последовательностей (мотивов):
1 - на расстоянии 27-30 п.н. левее точки начала транскрипции (+1) расположен ТАТА-мотив (ТАТА-бокс или Голдберга-Хогнесса-бокс). ТАТА-бокс включает последовательность, состоящую из 7 нуклеотидов:
5' – Т – А – Т – А – А – А – А – 3'
│ │ │ │ │ │ │
3' – А – Т – А – Т – Т – Т – Т - 5'
Он встречается у всех эукариотических генов (85%), поэтому его относят к неспецифическому сайту транскрипции. ТАТА-последовательность необходима для точного выбора кодирующей цепи ДНК и точки начала транскрипции м-РНК, в синтезе которых, принимает участие РНК-полиме-
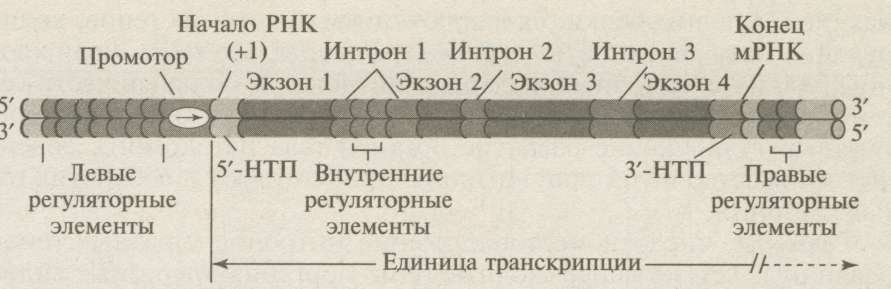
Рис.18. Структура эукариотического гена, кодирующего белок: 1+-точка инициации; 5′- НТП и 3′-НТП - 5′- и 3′-нетранслируемые последовательности.
раза II. В области расположения ТАТА-бокса происходит присоединение РНК-полимеразы к промотору, при участии комплекса белков - общих факторов транскрипции.
2 - на расстоянии 80-50 н.п. от точки начала транскрипции располо- жена последовательность ЦААТ (ЦААТ-бокс), характерная для тканеспецифических генов (глобинов, актина и др.) и регулирующая час- тоту инициации.
3 - на расстоянии 90 н.п. расположена последовательность ГЦ (5'- Г-Г-Г-Ц-Г-Г-3'), которая может встречаться в промоторе несколько раз и характерна для конституционных генов, обеспечивающих общеклеточные функции и влияние на скорость транскрипции.
Промоторы генов, адаптивных белков, образование которых усиливается под влиянием факторов среды включают дополнительные короткие последовательности, которые определяют транскрипцию этих генов только при наличии этих факторов: у белков теплового шока это последовательность -ЦТЦТ-, для генов металлсодержащих белков ею яв-ляется последовательность -ТГЦГЦТТГГ- и т.д. С этими сайтами связы- ваются специфические белки, регулирующие функции этих генов.
Терминаторами транскрипции генов эукариот являются ГЦ-богатые участки в конце генов. Сила взаимодействия внутри ГЦ-пар настолько сильна, что расплетание ДНК на этом участке становится затрудни- тельным. Это замедляет движение РНК-полимеразы и служит сигналом окончания транскрипции.
К регуляторной части эукариотического гена относятся особые последовательности – энхансеры (усилители)(см.выше). Они могут находится как на 5'-конце, так и на 3'-конце фрагмента ДНК, включающего ген, а также в составе интронов и действовать на больших расстояниях (более 1000 н.п.) через регуляторные белки, усиливая транскрипцию. Противоположную регуляцию осуществляют сайленсеры – последовательности нуклеотидов, имеющие такое же расположение на молекуле ДНК, но обеспечивающие ослабление транскрипции.
Кодирующая часть гена эукариот в составе транскриптона содержит:
1 - кодирующие последовательности экзоны и некодирующие после- довательности интроны (для мРНК);
2 - кодирующую последовательность и спейсеры (для рРНК), коди- рующую последовательность и один интрон (для тРНК). Интроны и спейсеры удаляются в ходе процессинга пре-РНК;
3 - 5'- и 3'-нетранслируемые последовательности (5'-НТП и 3'-НТП).
Экзон-интронную организацию имеют все гены, кодирующие белки позвоночных, за исключением генов гистонов и интерферонов. Экзоны имеют, как правило, небольшую длину. Длина интрона может быть разной – от несколько десятков пар нуклеотидов до многих тысяч. Общая длина интронов значительно превышает суммарную длину экзонов. В геноме человека только 1-2 % ДНК приходится на экзоны и 24 % - на интроны. Располагаются интроны в гене между сегментами, которые кодирует структурные или функциональные домены белка и характерна для каждого гена. В процессе транскрипции интроны считываются вместе с экзонами, а затем вырезаются в ходе процессинга (созревания) мРНК.
Для эукариот характерны моноцистронные единицы транскрипции и образуется мРНК, кодирующая только один белок.
Оперонный тип расположения генов, то есть, объединение генов в блоки под общим контролем регуляции для эукариот не характерен. Гены эукариот, контролирующие одну цепь биохимических реакции могут располагаться в разных районах хромосомы и даже в разных хромосомах. Гены эукариот чаще всего являются уникальными, встречающимися в геноме в единственным числе, но могут быть и многократно повторя- ющимися. Для некоторых генов известна кластерная организация. В кластер объединены гены, которые являются членами мультигенных семейств, которые содержат гомологичные последовательности одного гена -предшественника и их генные продукты часто имеют сходные функции. Белки, которые кодируется такими генами, называются паралогами.