

5.1.4. Репликация теломерных отделов днк
Теломеры (от греч. telos - конец) – это участки ДНК на концах плеч хромосом. Наличие на концах хромосом особых последовательностей ДНК впервые обнаружил Г.Мёллер в 1932 году.
Нуклеотидная последовательность теломерных отделов ДНК представляет собой последовательности, не несущие генетической инфор -мации:
Ц(цепь) 5′)ЦТААЦЦ-......-ЦТААЦЦ-.....-ГГТТАГ-....... - ГГГТАГ(3/)
Г(цепь) (3′)ГАТТГГ-...... .-ГАТТГГ-.......ЦЦААТЦ-.......-ЦЦААТЦ(5/)
Эта последовательность характеризуется наличием многочисленных повторов последовательности –ГАТТГГ- на 3′-концах каждой цепи ДНК. В каждой теломерной области ДНК различают Г- и Ц-цепь. Одна и та же цепь ДНК в одной теломерной области выступает как Г-цепь, в другой как Ц-цепь. Всего теломерные отделы ДНК включают тысячи указанных повторов. Их протяженность на одном конце составляет в клетках эмбриона человека 10-15 тыс. н.п. Таким образом, на обе теломерные области приходится 0,02% от средней длины молекулы ДНК человека (120 млн. н.п.). У мышей этот процент выше.
В области теломер более высокие уровни организации образуются за счет специфических белков. Если в основной части ДНК упаковывается на нуклеосомы, состоящие из гистоновых белков, то в теломерной области вместо гистонов в состав нуклеосом входят белки Rap1 и TRF1 (у млекопитающих). Благодаря этим белкам теломеры образуют более плотную упаковку, то есть относятся к гетерохроматину. Такая структура делает теломеры более стабильными (недоступными для действия ферментов – теломераз, ДНК-метилаз и эндонуклеаз (об их функциях речь пойдет ниже). Кроме этого, эти белки обеспечивают прикрепление теломер к компонентам ядерного матрикса.
Теломерная ДНК образует петли (в виде «лепестков ромашки»), фиксируемые на матриксе. По мере укорочения теломер число «лепестков» уменьшается. Укорочение теломер является результатом концевой недорепликации, имеющей место в процессе репликации ДНК. Впервые эту проблему сформулировал А.М.Оловников в 1971 году. (проблема касается только линейных ДНК).
ДНК-полимеразная система оставляет недореплицированными 3'-концы материнской ДНК, т.е., новые цепи оказываются укороченными с 5'-концов(рис.97).
Это происходит потому, что после удаления РНК-затравки специальной нуклеазой, образовавшаяся брешь не может застроиться, так как ДНК-полимераза способна только удлинять 3′-концы уже имеющегося полинуклеотида. Поэтому все бреши, возникающие после удаления РНК-затравки, заполняются; на конце хромосомы (β-брешь на рисунке) этого не происходит, так как, у этой бреши отсутствует следующий 3'-конец ДНК. Подобные концы ДНК (где одна цепь длиннее другой) называются острыми или оверхенгами. Острые концы атакуются экзонуклеазами, которые понуклеотидно отрезают одноцепочные участки. Концы ДНК становятся «тупыми». Укороченные молекулы ДНК составляют 50-60 н.п. (10-12 н.п. РНК-затравки и размеры ДНК-полимеразного комплекса), то есть укорочение ДНК за одно деление клетки составляет 0,00005% и через n количество делений хромосомы исчезли бы вообще.
Теломерные отделы ДНК, как было сказано выше, не несут генетической информации. Потому процесс укорочения теломер предохраняет от недорепликации более значительные области ДНК, следовательно, выполняет роль своеобразного буфера.
Теломерные участки выполняют также ряд важных функций, поэтому их укорочение допустимо лишь до некоторого предела. Поэтому в клетках происходит достраивание недореплицированных участков несколькими способами. Основной способ известный для всех клеток связан с действием теломеразы.
Теломераза является ферментом, который согласно номенклатуре, называется теломерной терминальной трансферазой. Она способна достраивать 3′-концы линейных молекул ДНК хромосом краткими (6-8 нуклеотидов), повторяющимися последовательностями (у позвоночных ТТАГГГ). Помимо белковой части, теломераза содержить РНК, которая выполняет функцию матрицы для достраивания ДНК. Длина теломеразной РНК у человека составляет 450 нуклеотидов. Благодаря наличию в теломеразе РНК, её следует считать специфической обратной транскрип- тазой, ферментом, обеспечивающем синтез ДНК на матрице РНК. Теломераза удлиняет не новую укороченную цепь ДНК , а старую – более длинную. Это происходит до того, как экзонуклеазы успевают «подравнять»
старую цепь под размер новой (рис.98). К 3′-концу старой цепи теломераза пристраивает несколько десятков или сотен повторов – ГГТТАТ, то есть удлиняет Г-цепь той или иной теломеры. Удлинение Г-цепи происходит за счет особенностей строения теломеразы. Средний короткий участок теломерной ДНК комплиментарен 1,5 теломерным последовательностям (-АУЦ-ЦЦА-АУЦ--). Триплет –АУЦ- связывается путем гибридизации с крайним теломерным полуповтором Г-цепи ДНК (ТАГ), остальная часть РНК служит матрицей для удлинения 3′-конца Г-цепи на 1 теломерный повтор. Этот этап называется элонгация теломеры. Затем происходит транслокация – перемещение фермента теломеразы вместе с РНК по удлинямой ДНК на 1 повтор левее, и опять продолжается элонгация. Таким образом пристраивается десятки и сотни повторов. В случае, если теломерная РНК мутирует, то теломераза теряет свою активность.
После этого, значительно удлиненная Г-цепь становится матрицей для образования фрагмента Оказаки на новой цепи так же, как это происходит на запаздывающей цепи. Для этого, в районе 3′-конца старой цепи праймаза синтезирует РНК-затравку, затем ДНК-полимераза β последовательно


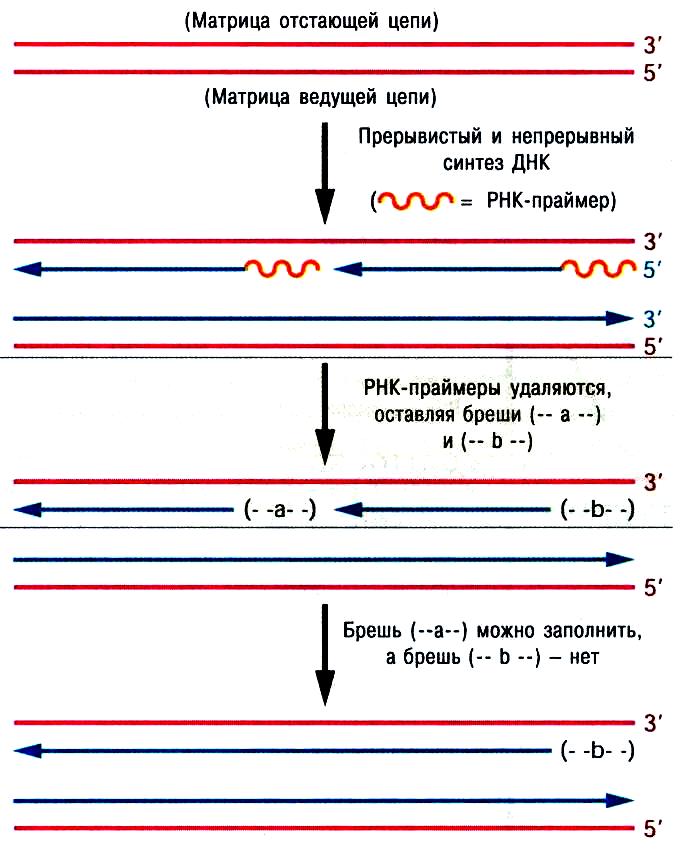
Рис.97. Недорепликация концов линейных хромосом
(Из: Клаг, 2009).
присоединяет комплементарно нуклеотиды родительских цепей. Рост цепи идет в направлении 5′→3′ и прекращается при достижении 5′-конца дочерней цепи. Сшивание фрагментов обеспечивает ДНК-лигаза. Затем экзонуклеаза удаляет РНК-затравку. Таков основной способ восстановления теломер. Новая цепь короче старой немного (10-15 нуклеотидов), а двуцепочная ДНК становится длиннее на серию теломерных повторов.
Впервые теломераза была открыта 1985 году у одноклеточных эукариот. В настоящее время с наибольшим постоянством она обнаруживается в органах кроветворения, в клетках периферической крови (гранулоциты, лейкоциты), эпителии кожи и слизистых оболочек, в эндометрии и др. В клетках, не способных делится, она не встречается. То
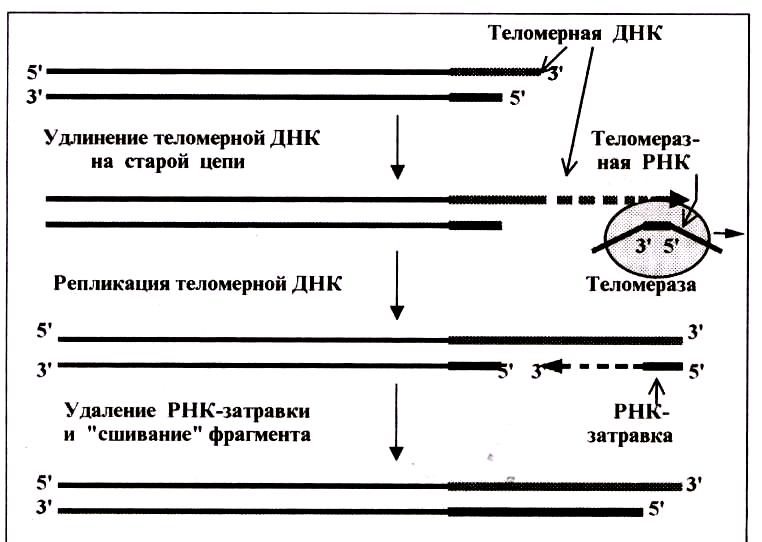

Рис.98. Удлинение теломерного участка ДНК с помощью теломеразы.
есть, теломераза обнаруживается только в клетках, способных к делению. Но наличие теломеразы не обеспечивает полного устранения явления укорочения теломер. По мере увеличения количества делений или увеличения возраста животного теломеразная активность снижается.
Кроме этого способа существуют альтернативные механизмы удлинения теломер (ALT – Alternative Lengthening of Telomeres). Они реализуются без действия теломеразы и известны для клеток дрозофил и линий некоторых опухолевых клеток. Один из способов – это рекомбинация между теломерными участками разных хромосом (рис.99). Суть способа заключается в том, что две молекулы ДНК взаимодействуют между теломерами, образуя гибридные теломеры. Более длинная цепь служит матрицей, по которой ДНК-полимеразы достраивают более короткие цепи. По окончании процесса либо восстанавливается структура теломер, либо молекулы ДНК обмениваются теломерными фрагментами. Таким образом, в не мейотической клетке происходит в ограниченной форме явление кроссинговера.
Восстановления теломерных отделов ДНК обеспечивает выполнение теломерами следующих функций:
а) теломеры участвуют в фиксации хромосом к ядерному матриксу, что обеспечивает правильную ориентацию хромосом в ядре (что особенно важно при кроссинговере в мейоза);
б) теломеры сцепляют друг с другом концы сестринских хроматид (после S-фазы интерфазы), возможно, за счет гидридизации теломер сестрин-ских хроматид. Если происходит мутация гена теломеразной РНК, расхождение хроматид блокируется;
в) наличие теломер предохраняет генетически значимые отделы ДНК от недорепликации, если в клетке отсутствует теломераза (или не выполняются ALT);
г) если есть теломера, то она обеспечивает стабилизацию концов разорванных хромосом, возникающих при разрыве хромосом. В отсутствии теломеразы фрагменты хромосом сливаются, деградируют, что блокирует клеточный цикл и ведет клетку к гибели. Данный феномен обусловливает возникновение α-талассемии при разрыве хромосомы 16q. Теломера же стабилизирует эти фрагменты и позволяет им функционировать.
д) теломеры влияют на экспрессию генов. Гены расположенные возле теломер репрессированы (эффект положения). Этот эффект называется сайленсинг или транскрипционное молчание. При укорочении теломер свыше допустимых пределов прителомерные гены активируются. Эффект положения может коснуться и внутренних генов, если они становятся транспозонами (генами, способными перемещаться в другой участок ДНК) и встраиваются в прителомерную область;
е) «счетная» функция теломер. Теломеры выступают в качестве часового устройства (репликометра), которое отсчитывает количество делений клетки после исчезновения теломеразной активности.
Таким образом, при достижении критически короткой длины теломеры теряют способность выполнять вышеперечисленные функции, что является одной из причин остановки клеточного цикла и гибели клетки.
Проблема концевой недорепликации теломер и роль теломеразы имеет большое биологическое значение. В частности, она напрямую связана с такими проблемами как старение и онкогенез.
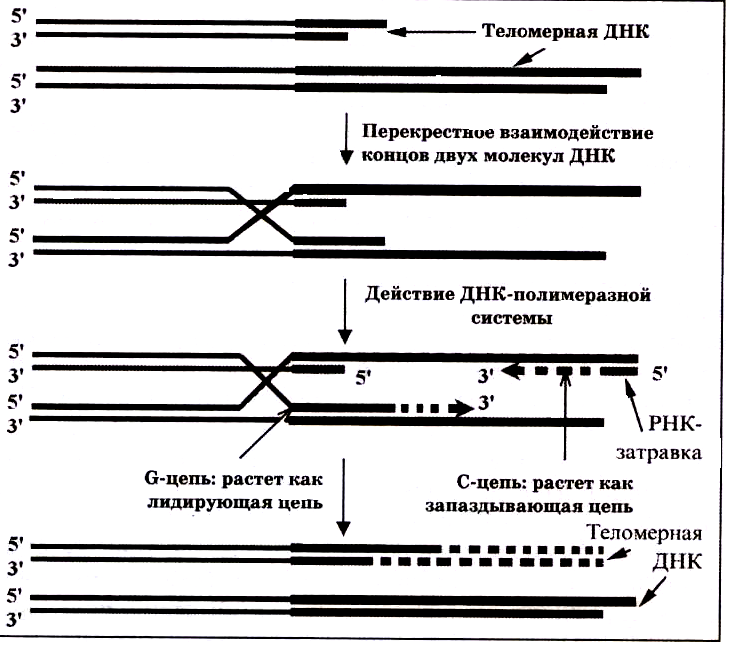

 Рис.99.
Удлинение теломерных участков
ДНК
путем рекомбинации.
Рис.99.
Удлинение теломерных участков
ДНК
путем рекомбинации.