

Раздел I. Организация генетического материала
Как было сказано выше, генетическим материалом клетки является молекула ДНК. Выделяют генный, хромосомный и геномный уровни организации генетического материала. Каждый уровень характеризуется своими свойствами, структурой и закономерностями передачи в ряду поколений.
Глава 1. Генный уровень организации генетического материала
Элементарной единицей функционирования генного уровня организации генетического материала и единицей генетической информации является ген. Ген – это участок ДНК, на котором закодирована информация о структуре РНК или полипептидной цепи. Последовательность нуклеотидов гена служит матрицей для синтеза разных видов РНК. В свою очередь мРНК служит матрицей для построения полипептидной цепи; последовательность аминокислот полипептида колинеарна последовательности кодонов гена. Белки, состоящие из одной или нескольких цепей, могут играть регуляторную или структурную роль (белки мембран), служить рецепторами для других молекул, выполнять транспортную функцию (гемоглобины), каталитировать определенную метаболическую реакцию (ферменты). Таким образом, свойство белка, определяемое последовательностью аминокислот, является элементарным или простым признаком. Признак – это единица морфологических, физиологических, биохимических, иммунологических и других качеств или свойств, по которым один организм отличается от другого. Следовательно, ген, кодируя последовательность аминокислот в белках, несет инфор- мацию о признаках данного организма. Передача генов в ряду поколений клеток или организмов обеспечивает наследование потомками признаков родителей.
Следовательно, гены – это участки ДНК, обеспечивающие контроль развития определенных признаков организма, управляющие этим развитием и всей жизнедеятельностью организма. Они контролирует клеточный метаболизм, благодаря содержащейся в них информации о структуре белков; белки-ферменты управляют всеми химическими процессами, являясь их катализаторами. Таким образом, ген – это единица наследственной информации, занимающая определенное положение в хромосоме или геноме и контролирующее выполнение определенной функции в организме.
В процессе изучения гена представление о его структуре, свойствах и действии менялось.
Первые предположения об организации наследственного материала были высказаны в 60-х годах ХIХ века основоположником генетики Г.Менделем. На основании своих опытов и данных предшественников (О.Сажре, Ш.Нодэн, Т.Найт, П.Люка) он пришел к выводу, что наследст - венный материал дискретен, то есть, представлен отдельными наслед- ственными задатками (факторами), отвечающими за развитие определенных признаков организма. Работы Г.де Фриза, К.Корренса и Э.Чермака (1900) подтвердили представление о существовании наследственных факторов. В 1906 году У.Бэтсон предложил термин «генетика» (от лат. geneticos – относящийся к происхождению) для обозначения бурно развивающейся науки о наследственности и изменчивости.
В 1909 году В.Иогансен предложил термин «ген» для обозначения наследственных задатков. Немецкий биолог Т.Бовери и американец У.Сэттон (1903) высказали предположение о том, что ген расположен в хромосоме.
В 1910 году группа английского биолога Т.Моргана (А.Стертевант, К.Бриджес и Г.Меллер) начали эксперименты, которые к 20 годам ХХ века позволили сформулировать хромосомную теорию наследственности. Одним из главных пунктов теории являлось положение, что гены расположены в хромосоме линейно. Таким образом, в 20 годах ХХ века сложилось представление о том, что ген – это элементарная неделимая единица наследственности, которая контролирует развитие одного признака.
В1929 году А.С.Серебровский и Н.П.Дубинин, еще не зная природу гена, на основании своих исследований пришли к выводу о сложности организации гена и его делимости.
Впервые предположение о связи между генами и белками-ферментами выдвинул английский врач Арчибальд Гэррод в 1902 году. Это предположение подтвердили 1941 году Дж. Бидл и Э.Тэйтум. Они изучали биохимическую роль генов в опытах с хлебной плесенью. Изучая мутации, возникающие у плесневых грибков под воздействием рентгеновского облучения, они обнаружили, что генетическая мутация фермента, необходимого на определенной стадии метаболизма приводит к образо- ванию его нефункциональной формы. В результате этого дальнейшая цепь биохимических реакций прерывается. Исходя из этого, Бидл и Тэйтум, сформулировали гипотезу «один ген - один фермент»: каждый ген направляет синтез одного фермента. Так, для превращения продукта А в продукт С необходим продукт В; мутация гена В (рис.1) приведет к тому, что продукт А будет накапливаться, а продукт В и С не образуется вовсе. Позднее было установлено, что многие белки имеют более сложную структуру, в образовании которой принимают участие несколько пептидных цепей (например, гемоглобин состоит из 2-, и 2 β-цепей), каждая из которых кодируется собственным геном. На этом основании В.Ингрэм (1957) предложил уточнить формулировку гипотезы «один ген – один фермент» в виде «один ген – одна полипептидная цепь». Из этого следует, что ген – это фрагмент молекулы ДНК, кодирующий один белок или, в случае сложности структуры белка, одну полипептидную цепь. В 30-х годах ХХ века были высказаны предположение о химической природе гена (работы.К.Кольцова, Н.В.Тимофеева-Ресовского, М.Дельбрука и др.) как части макромолекулы.
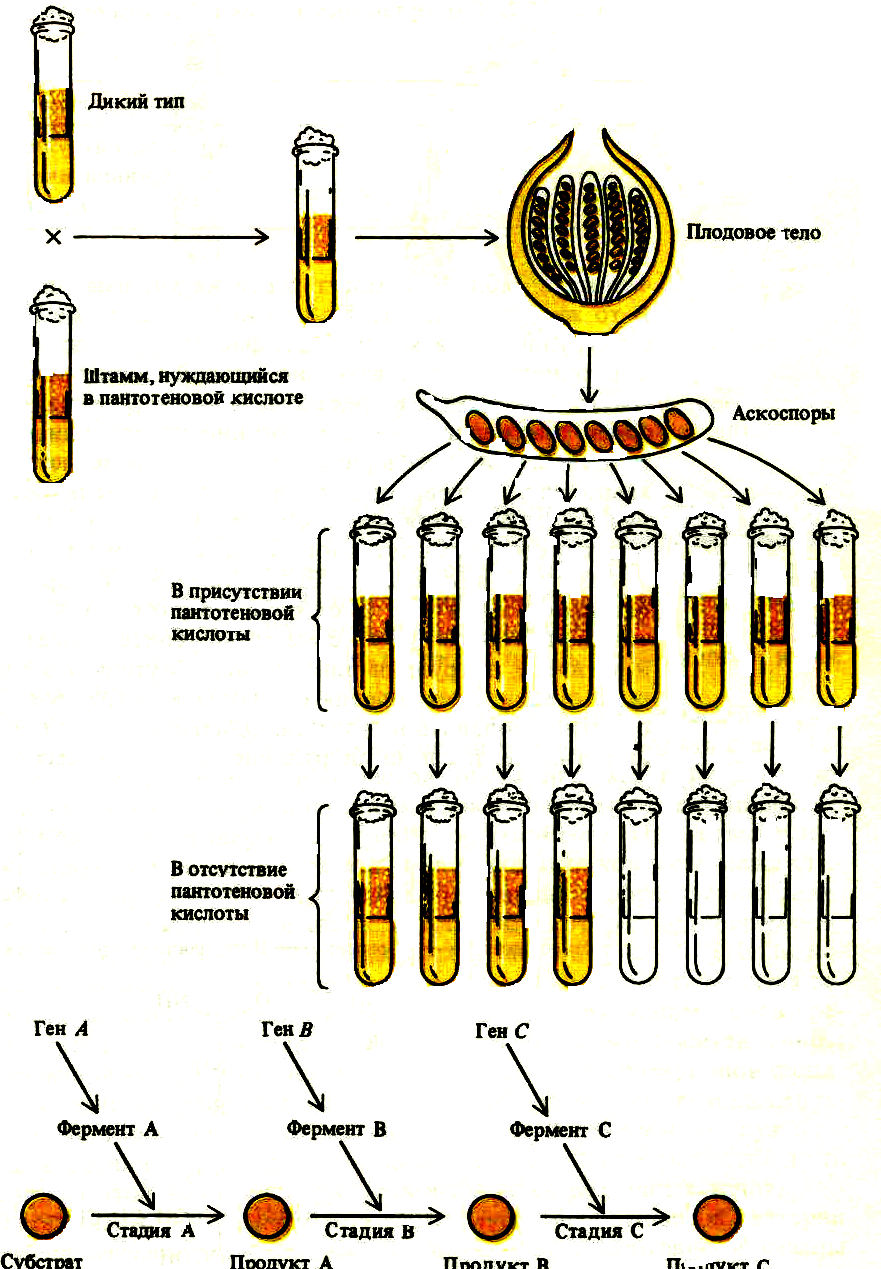
Рис.1. Схема иллюстрирующая гипотезу «один ген – один фермент»
(Из: Айяла-Кайгер,1988)
В 1944 году в результате работ по трансформации у бактерий О.Эверс, К.Мак-Леод и М.Мак-Карти доказали, что носителем наследственной информации является молекула ДНК, а в 1953 году была разработана модель молекулы ДНК Дж. Уотсоном Ф.Криком. С этого момента началось изучение тонкой структуры гена, как участка молекулы ДНК, несущего информацию о структуре полипептида.
Тонкую структуру ген изучал С.Бензер у бактериофага Т4. Им было установлено, что единицей функции являлось часть хромосомы – цистрон, мутации - мутон, рекомбинации – рекон.
Под термином цистрон понимают участок молекулы ДНК, несущий информацию о структуре одной полипептидной цепи. Если белок состоит из нескольких субъединиц (гемоглобин), то его ген состоит из нескольких цистронов (рис.2). У эукариот цистроны могут находиться рядом или локализоваться в разных хромосомах и обычно называются также генами (гены -цепи белка гемоглобина, ген -цепи белка гемоглобина). В настоя- щее время термины «ген» и «цистрон» используются как синонимы, но с генетической точки зрения термин «цистрон» является более точным для определения участка ДНК, кодирующего одну полипептидную цепь.
Р.Робетсон и Ф.Шарп с сотрудниками установили факт существо- вания расщепленных генов, то есть, генов, состоящих из кодирующих и некодирующих последовательности нуклеотидов, которые были названы, соответственно, экзонами и интронами. Экзоны – это кодирующие после- довательности нуклеотидов, а интроны – не кодирующие последовате- льности (рис.3). Интрон-экзонная структура гена характерна для генов эукариот, у которых количество интронов в гене может достигать 90% гена. У прокариот экзон-интронная организация гена практически отсутствует.
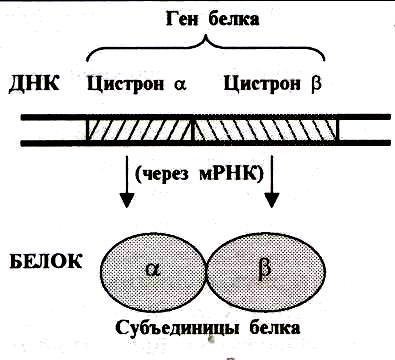



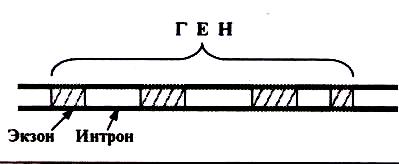


 Рис.3.
Экзон – интронная структура
Рис.3.
Экзон – интронная структура
гена.
Рис.2. Соотношение между геном
и цистроном.
Более поздние исследования показали, что неделимость гена в функциональном отношении не является абсолютным свойством.
Одна и та же последовательность нуклеотидов может обеспечить синтез нескольких полипептидов с разными функциями. Так, ген box митохондрий дрожжей, кодирующий дыхательный фермент цитохром в может существовать в двух формах: « длинный» ген содержит 6 экзонов и 5 интронов. Удаление из него 3 интронов приводит к образованию «короткого» гена белка РНК-матуразы. Этот белок обеспечивает вырезание остальных интронов и образование матрицы для цитохрома в. Это явление получило название альтернативного сплайсинга. Для вирусов и бактерий описаны случаи, когда один ген является частью другого или одна часть ДНК является составной частью двух генов. Такие гены называются перекрываю- щимися и для прокариот обеспечивают возможность синтеза большого количества белков при малых объемах генома. У эукариот также обнару- жены перекрывающиеся гены.
Делимость гена является так же результатом кроссинговера. Кроссинговер – это обмен наследственным материалом между гомоло- гичными хромосомами в процессе мейоза. Обмену предшествуют:
1 - образование хиазм (перемычек) между гомологичными хромо- сомами;
2 - разрыв хромосом в месте образования хиазм и обратное воссоединение;
3 - обмен наследственным материалом между гомологичными хромосомами в процессе их воссоединения после разрыва;
4 - образование новых сочетаний генов - рекомбинация генов.
Разрыв генетического материала может произойти внутри гена при неравномерном кроссинговере, что приводит к возникновению генной мутации (рис.4) по типу вставок и выпадений нуклеотидов. При этом наблюдается нарушение рекомбинации между хромосомами и искажение наследственной информации, но также происходит перераспределение нуклеотидных последовательностей в геноме. Совокупность этих процессов называется генетической рекомбинацией. Эти процессы лежат в основе генетической изменчивости. Наименьшая часть гена, рекомбинирующаяся в процессе кроссинговера, называется рекон и она является элементарной единицей рекомбинации.

-
1

2
3
4
5
6
7
8
9



 I
I

-
1
2
3
4
5
6
7
8
9



-
1
2
5
6
7
8
9


 II
II
-
1

2
3

4
3
4
5

6
7

8
9
III
Рис.4. Внутригенный кроссинговер с неравноценным обменом
наследственного материала : I – неравноценный крос -
синговер между гомологичными хромосомами; II – выпа-
дение 3 и 4 фрагментов; III – удвоение 3 и 4 фрагментов
В процессе мутации гена происходит изменение его химической структуры. Эти изменения являются результатом выпадения, вставки или замены нуклеотидов гена. В итоге наблюдается изменение последователь- ности нуклеотидов, которое приводит к появлению новых вариантов генов и, следовательно, новых признаков. Минимальное количество наследствен- ного материала, способного, изменяясь, приводит к появлению новых признаков, называется мутон. Мутон является элементарной единицей мутации. Наименьшая величина мутона равна одной паре нуклеотидов.
Таким образом, единицей генетической функции является цистрон, единицей генетической рекомбинации гена – рекон, а единицей генетичес- кой изменчивости – мутон.
Изменения наследственного материала (мутации), возникающие спонтанно или под действием различных факторов приводит к тому, что один и тот же ген может существовать в нескольких вариантах, которые различаются по содержанию в них биологической информации. Каждый вариант гена определяет синтез белков с разными свойствами, и, следо- вательно, разными вариантами развития данного признака ( признак – цвет семян гороха, варианты признака – желтый, зеленый). Форму существования гена, которая может определять развитие определенного конкретного варианта данного признака, называют аллелью.
Фактически аллель – это разновидность нуклеотидной пары, находя- щейся в данной точке молекулы ДНК на участке данного гена. Следовательно, истинных аллелей по данной точке гена, возникших в результате мутации, может быть 4: АТ, ТА, ГЦ, ЦГ (один из них аллель дикого типа, т.е. наиболее часто встречающийся в природе). Если в гене мутируют разные нуклеотидные пары, то две такие формы одного гена называются псевдоаллелями (или псевдогены). Общее число истинных аллелей и псевдоаллелей возможное для данного гена равно 4n, где n – число пар нуклеотидов в данном гене.
Аллели
гена располагаются в одном локусе
(участке) хромосомы, который может
содержать только один из серии аллелей.
Это делает аллели альтернативными
(взаимоисключающими) вариантами
существующего гена. Присутствие в
генофонде вида (совокупность всех генов
данного вида) одновременно более двух
аллелей называется множественным
аллелизмом.
Например, у человека – это ген ,
определяющий группы крови системы АВО
и имеющий 3 аллеля (IА,
IВ,
IО);
более 100 аллелей насчитывают гены -
и β-глобулинов; множественный аллелизм
присущ генам иммуноглобулинов. тации
()вению генной мутации ()Рис. приводит
внутри гена)мами в процессе мейоза.