

2.2.2. Наднуклеосомная укладка днк
Дальнейшая спирализация хроматина приводит к образованию плотного тела митотической хромосомы и включает следующие уровни компактизации: хроматиновая фибрилла, хромомерный и хромонемный, хроматидный и метафазную хромосому.
Хроматиновая фибрилла. Этот уровень компактизации обеспечивается гистоном Н1, который будучи связанным с линкерной ДНК, связывается также с соседними нуклеосомами и, изменяя свою конформацию, сближает их (рис.32). При этом образуется глобула, из 6 нуклеосом ( сверхбусина), или соленоид (рис.30,b).
Такая структура называется элементарной хроматиновой фибриллой или нуклеомером; имеет диаметр 30нм и длину 1,2мм. Уровень компактизации составляет 40 раз (рис.34). На один нуклеомер приходится 1600 н.п.
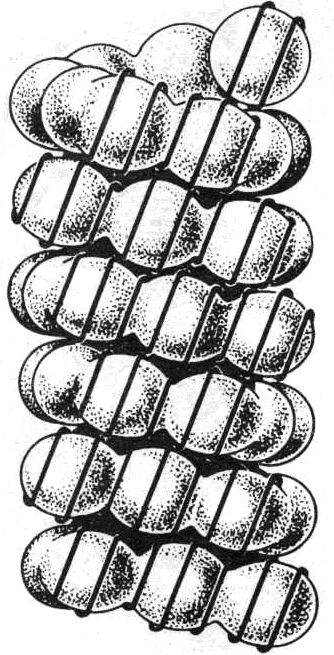
Рис.34. Элементарная хроматиновая
Фибрилла – нуклеомер (Из: Жимулев.2007).
Хромомеры и хромонемы. Следующий уровень структурной компактизации ДНК связан с укладкой хроматиновой фибриллы в петли (рис.35).
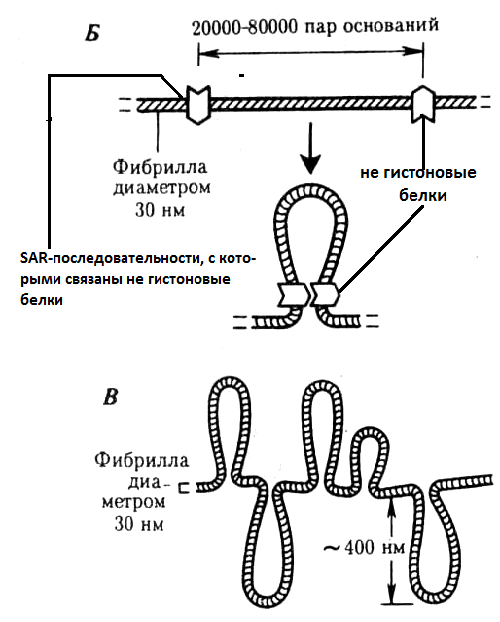 Рис.35.
Петлевой
уровень
компак-
Рис.35.
Петлевой
уровень
компак-
тизации хроматина
Соленоидная хроматиновая фибрилла складывается в петли разной длины. В образовании петель участвуют не гистоновые белки ядерного матрикса, которые способны узнавать специфические последовательности ДНК (SAR- последовательности), отдаленные друг от друга на расстоянии в несколько тысяч пар нуклеотидов. Эти белки образуют остов (scaf- fold), который связывается с SAR-последовательностями ДНК (scaffold attachment regions) (рис.36) и, сближая их, образует петли (рис.35). Каждая петля содержит от 20 до 80 тыс. пар нуклеотидов, что может соответс- твовать одному или нескольким генам и называется петлевой домен (рис.30,с). В интерфазном ядре такие петли связаны с сетчатым белковым образованием, расположенным на внутренней стороне ядерной оболочки и называемым сетчатым ядерным матриксом, ламиной или ядерной пластинкой (рис.37). Белки ядерного матрикса ламины соединяются с последовательностями ДНК, которые называются МАR-последовательностями (matrix attachment regions).
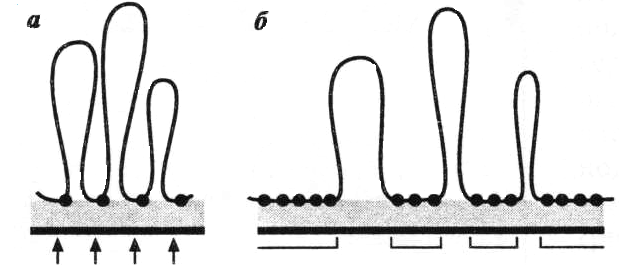
Рис.36. Модели закрепления оснований ДНК в ядерном матриксе (Razin 1995), (черными кружками обозначены МАR-последовательности; серая полоса-скаффолд, состоящий из белков ядерного матрикса. Из: Жимулев, 2007).


Рис.37.Схема ядерной мембраны (СЯМ – сетчатый ядерный матрикс)
(Из: Фаллер, 2003)
В результате такой спирализации хроматиновая фибрилла диаметром 30 нм преобразуется в структуру диаметром 300 нм, называемую интерфазной хромонемой. Отдельные участки хромонемы подвергаются дальнейшей компактизации : соседние петли объединяются в структурные блоки в виде розеток (рис.38), петли которых, сильно спирализуясь, образуют хромомеры, небольшие, сильно окрашивающиеся компактные тельца (0,1-0,20мкм), различающиеся по форме и размеру. Число и рисунок хромомер у данного организма на данной стадии клеточного цикла одинаковы и постоянны (рис.39).

Рис.38. Спирализация хромонем с об - разованием розетковидных участков: петли фибриллы диаметром 30 нм(А) соединяются в розетки (Б); в резуль- тате упаковки (спирализации) каж- дой петли розетки образуются ком -пактные тельца – хромомеры (В);
( Из: Захаров и др., 1982)

Рис. 92. Хромомеры хромосомы XII тритона: а – в нитях хромосом клеток Allium cepa; б) в нитях сестринских хроматид (черные точки)
Петлевые домены и хромомеры расположены неравномерно . Неодинаковая степень спирализации разных участков хромосом имеет большое значение; в зависимости от степени спирализации выделяют эухроматин и гетерохроматин.
Р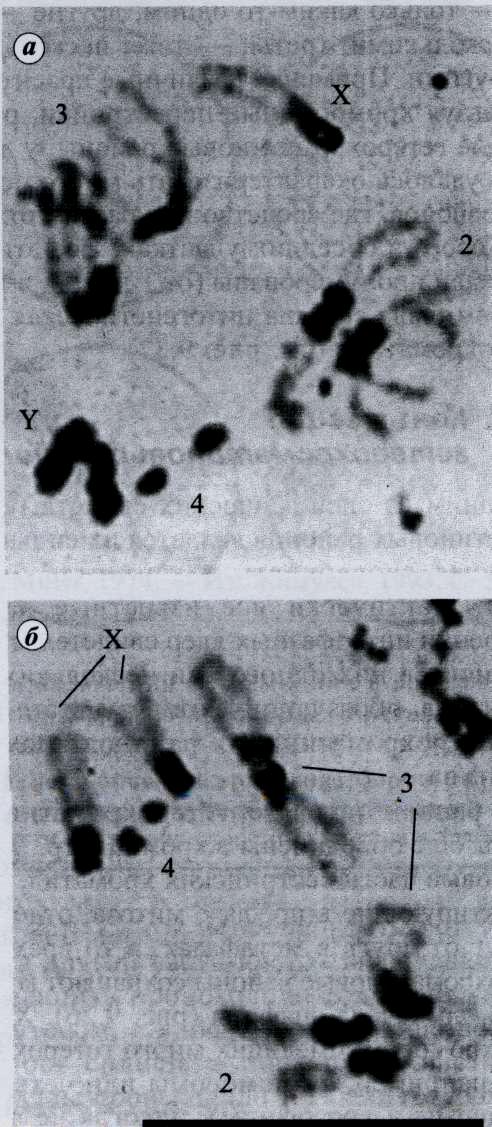 ис.40.
Эухроматин (светлые участки) и
гетерохроматин (темные участки) в
кариотипе дрозофилы (из Жимулев, 2007):
цифры – номера хромосом.
ис.40.
Эухроматин (светлые участки) и
гетерохроматин (темные участки) в
кариотипе дрозофилы (из Жимулев, 2007):
цифры – номера хромосом.
Эухроматин – это участки хроматина, имеющие меньшую степень спирализа- ции в неделящихся клетках; на препаратах выглядят как более светлые участки (рис.40). Эухроматин является активной областью хроматина, здесь постоянно идет транскрипция ДНК. На время митоза эти участки сильно спирализуются и транскрипция с них прекращается.
Гетерохроматин (рис.40) – это сильно спирализованные, компактно конденсированные, неактивные участки хроматина. Транскрипция биологической информации в их пределах не происходит. Чаще всего гетерохроматин располагается в прицентромерных, иногда в теломерных участках хромосом или, весьма редко, как вкрапления в эухроматине. Такой гетерохроматин называется интеркалярным. В световом микроскопе он виден в виде глыбок или гранул. Часть гетерохроматина прилежит к кариолемме (примембранный хроматин),часть сосредоточена вокруг ядрышек (перинуклеарный хроматин).
Различают два вида гетерохроматина: конституционный и факультативный.
Конституционный гетерохроматин – это хроматин, с которого ни в одной клетке никогда не идет считывания информации в виде мRНК. Расположен в прицентромерных и теломерных областях хромосомы. Его роль заключается:
1 - в поддержании структуры ядра;
2 - в прикреплении хроматина к ядерной оболочке;
3 - в узнавании гомологичных хромосом в мейозе;
4 - участие в в регуляции активности генов и разделении соседних генов.
Факультативный гетерохроматин – это участки хроматина, спирализация (конденсация) которых служит механизмом выключения из активной функции генов, транскрипция которых не требуется в клетках определенной специализа- ции. Его практически нет в эмбриональных клетках, по мере их диф- ференциации количество этого хроматина увеличивается. Другим примером является одна из двух Х-хромосом женского пола, которая плотно упакована в виде телец Бара (тельца полового хроматина). Гены этой хромосомы не транскрибируются.
Образование хромонемы и хромомер укорачивают длину ДНК в 1000 раз.
Петельно – доменная структура хроматина имеет диаметр 300 нм и является интерфазной хромосомой. Её дальнейшая спирализация ведёт к образованию хроматиды с диаметром 600-700 нм.
Последняя степень спирализации (в 7000 раз ) происходит в профазе митоза; в результате образуется метафазная хромосома с диаметром 1400нм. Укладка хроматиновой нити на этой ступени спирализации представляет собой её многократное складывание по длине хромосомы (рис.41). Этот процесс начинается в профазе митоза, достигает максимального выражения в метафазе. В телофазе митоза происходит декомпактизация (деспирализация) вещества хромосомам, что приводит к восстановлению интерфазного хроматина.
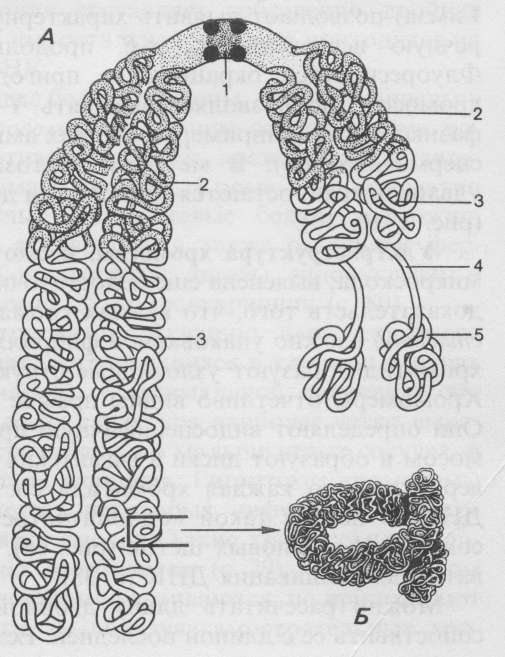
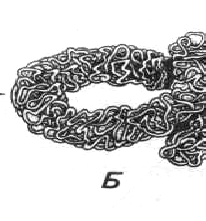
Рис. 94. Схема метафазной хромосомы (Из: Хадорн, 1988):
А) 1 - центромера с кинетохором; 2 – гетерохроматин; 3 – эухроматин;
4 - вторичная перетяжка; 5 – спутник;
Б) участок с более сильным увеличением (сверхспирализация)