

2.2.2.3. Микротрубочки
Микротрубочки (МТ) (рис.115) - это компонент цитоскелета, представляющий собой длинные нитевидные структуры, протянутые по всей цитоплазме и формирующие сеть, которая поддерживает структурную организацию и локализацию органелл.
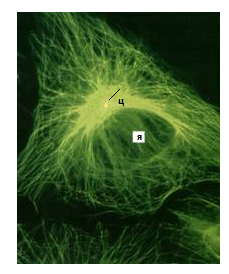
Рис. 115.Сеть микротрубочек, окрашенная мечеными антителами к тубулину
в клетке культуры ткани: в центре клетки желтым окрашена центриоль (ц); я - ядро (Из:Интернет-ресурсы – структура и функции цитоскелета)
Они играют важную роль:
1 - при делении клетки (116,Б);
2 - в процессе внутриклеточного транспорта, особенно, при перемешении пузырьков;
3 – в циркуляции веществ из комплекса Гольджи в ЭПС;
4 - в обеспечении подвижности клетки;
5 - в обеспечении структуры некоторых органелл (реснички, жгути - ки) (рис.116,А)
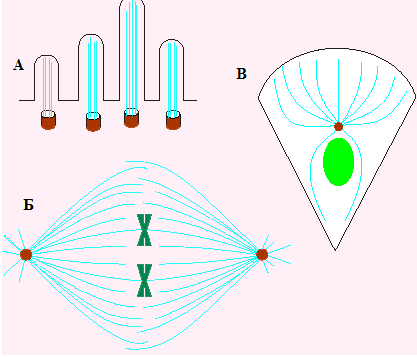
Рис.116. Формы участия микротрубочек в функционировании клетки: структурная роль в образовании ресничек (А), веретена деления (Б), цитоскелета клетки (В) (Из:Интер - нет-ресурсы – структура и функции цитоске - лета)
Микротрубочки являются органеллами фибриллярного типа и представляют собой полые цилиндры. Микротрубочки образованы белком тубулином, состоящим из двух субъединиц: α- и β-тубулина (см.выше, основные белки цитоскелета). Субъединицы тубулина образуют гетеродимеры, которые складываются в протофиламенты. 13 протофиламентов формируют стенку цилиндра микротрубочки (рис.106). Внешний диаметр полого цилиндра составляет 28 нм, внутренний – 15нм. Субъединицы тубулина располагаются в шахматном порядке. Протофиламенты соединяются в цилиндрическую закрученную структуру, соединяясь полярными частями субъединиц. От микротрубочек отходят, ассоциированные с ними, молекулы белков (ассоциированные с микротрубочками протеины или МАР). МАР стабилизирует микротрубочки, участвует в регуляции разборки микротрубочек, а также связывают их с другими элементами цитоскелета и органеллами. Особую группу МАР составляют микротубулярные двигатели (белки кинезины и динеины), которые перемещаются вдоль по трубочкам. МАР специфичны для каждого типа клеток. Так, для нервных клеток характерны высокомолекулярные МАР (масса 200-300 кДа): МАР-1 и МАР-2, а также тау-белки (молекулярная масса 55-65 кДа). Белки обоих классов имеют домен, связывающий микротрубочки с клеточными компонентами. Считается, что МАР предотвращают деполимеризацию и стабилизируют микротрубочки. Микротрубочки являются динамичными структурами (рис.117). Они имеют два конца: (-) и (+)-концы (рис.117,А). Плюс-конец может расти очень быстро за счет присоединения молекул тубулина (полимеризация микротрубочек); минус-конец теряет тубулиновые субъединицы (деполимеризуется) до тех пор, пока не стабилизируется. Стабилизация минус-конца микротрубочки достигается присоединением (заякориванием) минус-конца микротрубочки к центросоме (рис.117,Б), называемой также центром организации микротрубочек (ЦОМТ) или клеточным центром, локализованном в клетки рядом с ядром (рис.116,В). От ЦОМТ микротрубочки растут к периферии клетки. Микротрубочки могут внезапно прекратить свой рост и укоротиться обратно по направлению к центросоме вплоть до полного разрушения, а затем вновь начать расти.

Рис. 117.Микротрубочки: стабилизация и функции (Из:Интернет-ресурсы – структура и функции цитоскелета)
Сборка и разборка микротрубочек связана с затратами энергии ГТФ. МТ постоянно растут, разбираются, растут вновь, и в клетках их система сохраняет радиальное строение с центром в центросоме (рис.116,В). Если рост и разборка МТ, чередуясь, происходят то на одном конце (плюс-конце), то на другом (минус-конце) края клетки, то такая нестабильность называется динамической (рис.118А); если рост МТ происходит на плюс-конце, а разборка – на минус-конце, то такая нестабильность называется тредмиллингом (рис.118Б). В клетках, имеющих центросомы, преобладает динамическая нестабильность микротрубочек, а в отсутствие центросомы - тредмиллинг, который приводит к снижению количества полимеризован – ного тубулина. Это доказывает роль центросомы в стабилизации минус-концов микротрубочек. Динамическая нестабильность микротрубочек играет важную физиологическую роль. Например, при делении клетки микротрубочки растут очень быстро, что способствует правильной ориентации хромосом и образованию митотического веретена деления.
Таким образом, ЦОМТ постоянно производит микротрубочки: одни микротрубочки растут от центра клетки к периферии, другие сокраща- ются за счет потери тубулиновых субъединиц. Сборка микротрубочек происходит в присутствии ГТФ, ионов Мg и температуре 37ºС (это условия полимеризации тубулинов) и проходит три фазы:
1 - нуклеация - замедленная фаза - формируется ядро из субъеди- ниц тубулина;
2 - элонгация - фаза полимеризации - ядро тубулина быстро удли-няется за счет свободных молекул тубулина. Полимеризация идет быстрее, чем деполяризация и микротрубочки быстро растут.

 3
- фаза
стабильного состояния
- поляризация и деполяризация урав-
новешивают друг друга.
3
- фаза
стабильного состояния
- поляризация и деполяризация урав-
новешивают друг друга.
Рис.118. Динамическая нестабильность (А) и тредмилинг (Б) микротрубочек
Движение микротрубочек и перемещение частиц по микротрубочкам обеспечивают молекулярныe двигатели - семейство моторных белков, участвующих в транспорте органелл, митозе и мейозе. К этим белком относятся динеины, кинезины и миозины I и II, которые взаимодействуют со многими компонентами цитоскелета (табл.20). Динеин и кинезин одним концом прикрепляются сбоку к микротрубочке и могут двигаться по ней, если доставлять им энергию в виде АТФ. При этом большинство вариантов кинезина двигается по трубочке к ее плюс-концу, а все динеины - к минус-концу. Другим полюсом молекула динеина или кинезина может прикрепиться к мембранным органеллам (рис.114) или к другим микротрубочкам (рис.119). Соединяясь с органеллами, микротрубочковые моторы перемещают их в клетке. Направление движения органелл будет разным в зависимости от того, какой мотор работает: кинезин "повезет" органеллу к плюс-концу микротрубочки, то есть к периферии, а динеин - к центру клетки. В жгутике и ресничке молекулы динеина, прикрепляясь к двум соседним микротрубочкам, заставляют эти микротрубочки скользить относительно друг друга (рис.119а,б). Если микротрубочки своими основаниями (минус-концами) закреплены на базальных тельцах, то скольжение соседних микротрубочек ведет к тому, что жгутик изгибается (рис.119,в).
Кроме микротрубочковых моторов, существуют еще миозиновые молекулы, способные перевозить органеллы вдоль актиновых микро- филаментов (см.выше). Какое различие между этими двумя видами транспорта в клетке? Установлено, что микротрубочки, идущие на дальние расстояния, подобны «скоростным шоссе», по которым органеллы быстро перемещаются между центром и периферией клетки. Микрофиламенты играют роль местных дорог. Доехав по микротрубочке до нужной области клетки, органелла меняет мотор и пересев на местный микрофиламент доезжает точно до места назначения. Микротрубочки и микрофиламенты, могут, в свою очередь, быстро менять свою длину и положение в результате полимеризации-деполимеризации, что также обеспечивает ускорение и точность доставки. Таким образом, органеллы могут передвигаться по клетке по одним и тем же рельсам-микротрубочкам в разных направлениях, пользуясь разными молекулами –моторами
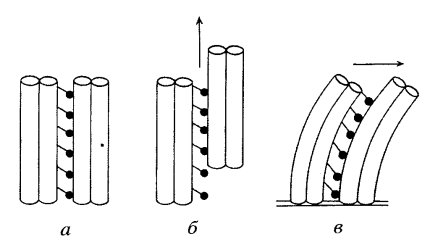
Рис.119. Смещение дублетов МТ при их движении:а - дублеты незакрепленных МТ;б - они же при смеще- нии относительно друг друга (скольжении); в – из- гибание дублетов МТ при закрепленных концах микро- трубочек (изгибание ресничек и жгутиков) (Из:Интернет-ресур-
сы – структура и функции цитоскелета)
.
Функции микротрубочек:
1 - играют роль цитоскелета;
2 - участвуют в транспорте веществ и органелл в клетке;
3 - участвуют в образовании веретена деления и обеспечивают рас- хождение хромосом в митозе и мейоз;
4 - входят в состав центриолей, ресничек, жгутиков;
5 - стабилизируют форму клетки.
Если клетки обработать колхицином, разрушающим микротрубочки, то клетки изменяют форму, сжимаются, теряют способность делиться. Колхицин связывается со свободными мономерами тубулина, которые вследствие этого теряют способность вставляться в растущие микротрубочки. Рост ее с плюс-конца останавливается, а разрушение с минус- конца продолжается вплоть до полного разрушения микротрубочки. Таким же свойством обладают белки винбластин и винкристин. Это явление используется в медикаментозной терапии онкологических больных. Антимитотические лекарственные вещества препятствуют образованию митотического веретена деления, останавливая быстрое деление клеток и рост опухолей.
Таблица20
 Молекулярные
двигатели в эукариотической клетке
Молекулярные
двигатели в эукариотической клетке