

Глава 3. Вертикальні структури геосистеми: міжелементні відношення та процеси (процесна ландшафтна екологія)
§ 3.1. Генетико-еволюційні відношення
Коротка історична довідка. Найбагатші традиції генетико-ево- люційного аналізу природних систем були в російській, а згодом у радянській фізичній географії. В. В. Докучаев та його учні (Г. Ф. Морозов, О. М. Краснов, І. М. Крашенінніков та Ін.) надавали генетичним аспектам дослідження природи надзвичайної ваги, тому докучаївську географічну школу небезпідставно називають генетичною. Та якщо в грунтознавстві генетичний принцип дійсно послідовно дотримувався і розвивався, то в ландшафтознавстві багато в чому він був декларативним, на практиці спрощувався. Так, часто дослідження генетичної суті ландшафту зводилось до аналізу його геолого-геоморфологічної будови, яка буцімто генетично зумовлює всю його структуру. Такий підхід особливо яскраво виявився в працях М. А. Солнцева та прихильників його наукового напряму.
У класичному ландшафтознавстві генетичний підхід визнавався як провідний при виділенні ПТК усіх рангів, їх типології, при районуванні, вирішенні інших питань. Проте, незважаючи на гіпертрофовано велику увагу ландшафтознавців до генетико-еволюцій- них проблем, їх грунтовно розроблено не було. Це зумовило появу в 70-х роках критичних статей щодо генетичного принципу та його евристичного й практичного значення (Д. Л. Арманд, Ф. М. Мільков та ін.). Було наголошено, що генетичні зв’язки між геокомпо- нентами аж ніяк не вичерпують усієї багатогранності внутрішньогеосистемних відношень, багато з яких практично незалежні від походження та особливостей розвитку геосистеми.
В екології спроби розв’язання питань генезису та еволюції екосистем були пов’язані з залученням до аналізу екосистем класичних дарвінівських конструкцій та методів біогеографії, проте важливих результатів це не дало. Так само не слід вважати за вдалу спробу отримати ці результати на основі концепцій екології, що намагався зробити Е. Піанка у своїй монографії «Еволюційна екологія» (1976). Дійсно еволюційним аспектам у ній відведено надто мало місця, а самі результати виявилися тривіальними.
Розв’язання цієї важливої проблеми в рамках класичного ландшафтознавства та екології наштовхнулось на труднощі і потребує ширшого залучення нових загальнотеоретичних положень (зокрема, інваріанту, нуль-моменту, інерційності, відносної автономності геокомпонентів тощо)..Значні перспективи в розвитку проблеми генетико-еволюційних відношень геосистем можуть уявлятися й широкому використанні результатів сучасного палеоландшафто- знавства, багато з останніх досягнень якого мають ландшафтно- екологічний зміст (Веклич, 1990).
Загальна схема та основні положення. Мета аналізу генетико- еволюційних відношень між складовими вертикальної структури геосистеми — визначити закономірності поєднання геокомпонентів один, з одним та взаємозалежність їх окремих властивостей. Він зводиться до вирішення двох основних груп питань: виявити закономірності формування даного поєднання геокомпонентів та характер зв'язків між ними; оцінити ступінь генетичної залежності між геокомпонентами та їх елементами.
Загальна схема формування вертикальної структури.геосистеми така. Утворення її починається з нуль-моменту—появи наземного твердого абіотичного субстрату. Цим моментом може бути вихід території з-під рівня моря, вивільнення її з-під льодовика, перекриття поверхні вулканічною лавою тощо. Геологічні породи, що вийшли на поверхню, грунтові води та приземний шар атмосфери вступають у взаємодію між собою, яка виражається у вивітрюванні гірських порід та зміні морфології рельєфу. Такі геосистеми мають примітивну вертикальну структуру, у якій немає біотичних компонентів. Проте вже на абіотичній стадії розвитку геосистеми до неї з атмосфери та з поверхневими водами надходять мікроорганізми, життєдіяльність яких спричиняє формування та поступове збільшення вмісту органічної речовини у верхньому шарі гірських порід. Внаслідок цього стає можливим ецезис (приживання на новому місці) лишайників, мохів, а згодом і вищих рослин, їх спори та насіння заносяться до геосистеми практично з нуль- моменту, але на безорганічному субстраті проростати вони не могли. Популяційна структура первинних рослинних угруповань примітивна і цілком визначається фізико-хімічними особливостями субстрату, а також видовим складом рослинності прилеглих геосистем. Конкурентні та інші біотичні відношення між популяціями та їх окремими особинами виражені слабко.
3 моменту виникнення фітоценозу геосистема вступає у біотичну: стадію розвитку. Для неї характерні інтенсифікація гумусо- утворення, заселення тваринами та формування зооценозу, більша Швидкість сукцесійних змін грунту, рослинності та мікробоценозів, Стабілізація рельєфоформуючих процесів, більша трансформація приземного Шару атмосфери (зокрема, внаслідок все більшої транспірації). Якщо на абіотичній стадії розвитку зміни геокомпо- нентів та їх зв’язок загалом детерміновані та односпрямовані, то на біотичний відношення між геокомпонентами значно ускладнюються і набувають все більшої стохастичності (ймовірносності). Формується розгалужена мережа обернених міжгеокомпонентних звязків, тобто переважно односпрямовані відношення змінюються двосторонніми. Так, розвиток грунту починає залежати нетільки від властивостей гірських порід, атмосфери і грунтових вод, а й від популяційного складу та продуктивності рослинних угруповань! У свою чергу, зміни грунту зумовлюють і зміни рослинності, тваринного населення (особливо грунтової фауни), мікробоценозів, які знову-таки зумовлюють певні зміни грунтових процесів. За таких зв’язків визначення геокомпоненту, який є провідним у формуванні вертикальної структури геосистеми, лишається сенсу;
Важлива особливість еволюційних відношень на біотичній стадії формування геосистеми зумовлена зростаючою самостійністю розвитку окремих геокомпонентів у процесі їх еволюції. Так, на перших етапах сукцесії рослинних угруповань формування їх популяційної структури майже повністю визначається абіотичними факторами — кліматом та субстратом. Ці фактори ніби задають генеральний напрям можливих змін рослинності. У подальшому роль внутрішньогеокомпонентних процесів (між- та внутрішньовидової конкуренції, симбіозу та ін.) набуває все більшого значення, і популяційний склад біоценозів все більше визначається власне цими відношеннями. Вони до певної міри узалежнюють фітоценоз від грунту, клімату та геологічної будови. Аналогічно і грунт у процесі свого розвитку стає все менш залежним від материнської породи та клімату і в рамках деякого заданого ними діапазону змін може розвиватися відносно самостійно. З розвитком грунтового профілю геологічні породи також стають менш залежними від атмосфери, рослинності, мікроорганізмів. Це дає змогу в одних кліматичних умовах на одному типі і навіть виді грунту формуватися різним рослинним угрупованням, різним видам грунту на одній геологічній породі тощо.
Таким чином, відповідність геокомпонентів один одному жорстко не детермінована. її характер можна визначити як детерміновано-стохастичний. Він полягає в тому, що вплив певного геокомпонента на інший (наприклад грунту на рослинність) визначає деякий діапазон змін рослинності, за який вони не можуть вийти, тоді як у рамках цього діапазону формування рослинних угруповань може визначатися суто внутрішньоценточними або й чисто випадковими процесами (наприклад, на дерново-підзолистих піщаних грунтах не можуть формуватися букові ліси, типчаково- ковилові степи, а різні асоціації хвойних та й мішаних лісів.тут можливі). Так само хоч і формування грунту обмежене характером рослинності, геологічною будовою, кліматом, рівнем та хімічним складом грунтових вод, ці обмеження не настільки значні, аби визначити єдино можливий вид грунту у даних умовах (на плоских лесових рівнинах при глибокому рівні залягання грунтових вод за однакових умов посушливого південно-степового клімату можуть формуватися чорноземи звичайні, різні, за вмістом гумусу, потужністю, глибиною залягання карбонатів і навіть, ме-

звужується (фільтрується). Послідовне врахування впливу всіх геокомпонентів на двосторонній зв’язок якихось двох з них приводить до виділення лише тих можливих варіантів поєднання геокомпонентів, яке може скластися в умовах конкретної геосистеми. Графічно приклад такого аналізу, що формально нагадує процес послідовної фільтрації зв’язків, зображено на рис. 5.
Ступінь генетичної залежності одного геокомпонента від іншого можна оцінити кількісно за допомогою поліхоричного показника зв’язку (синонім — коефіцієнт взаємоспряженості). Його значення змінюються від 1 (компоненти однозначно зв'язані між собою — кожному виду грунту відповідає лише певний один вид фітоценозу) до 0 (геокомпоненти не зв’язані між собою — на певному виді грунту можуть формуватися зовсім різні асоціації). Формули та техніку розрахунку поліхоричного показника зв’язку наведено в будь-якому підручнику з математичної статистики. Обчисливши значення цього показника для всіх пар геокомпонентів» отримаємо матрицю тісноти генетичних міжгеокомнонентних відношень, графічним зображенням якої може бути граф (рис, 6). Різні типи геосистем мають різний характер і силу генетичного
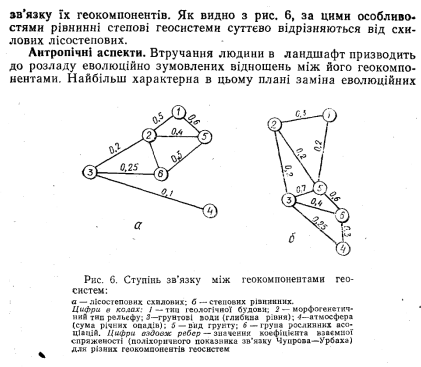
зв’язків між грунтом та рослинністю у природних геосистемах на антропічно регульовані відношення між ними в агрогеосистемах. Зведення лісів та їх заміна на трав’яні агроценози призводить до формування протиприродних, «еволюційно-абсурдних» зв’язків між грунтом та рослинністю, тваринним населенням, мікробоценозами. Наприклад, з природно-еволюційної точки зору зовсім нехарактерні відношення між дерново-підзолистими грунтами та злаковою сільськогосподарською рослинністю (еволюційно ці грунти пов"язані з мішаними або хвойними лісами). Якщо грунт як більш інерційний геокомпонент може тривалий час зберігати свої еволюційно- генетично зумовлені риси при взаємодії з новим біоценозом, то самі ці біоценози самостійно існувати в геосистемі не можуть, •еволюційно відштовхуються нею. Так, в Україні значні ареали сірих лісових грунтів постійно знаходяться під ріллею ще з XVI — XVII ст., проте свого генетичного типу вони не змінили. Натомість, існування агроценозів було можливим лише завдяки їх щорічному відновленню людиною. Припинення культивування агро- денозів приводить до поступового налагодження еволюційних відношень у геосистемі, і її корінний фітоценоз відновлюється (у лісових геосистемах у середньому через 300—600, в трав’яних — через 40—80 років).
Господарська діяльність людини призводить не тільки до видозміни еволюційних відношень між грунтом та біокомпонентами, а й викликає більш глибоку трансформацію внутрішньогеосистемних генетично зумовлених зв’язків, торкаючись водного режиму, рельє- фоформуючих процесів тощо. Так, у генетичному відношенні рельєф геосистем Причорноморської низовини акумулятивного походження, проте з масовим розоранням земель цього регіону, іригацією зараз тут переважають рельєфоутворюючі процеси, характерні не акумулятивним, а денудаційним рівнинам.
Надмірне намагання штучно законсервувати геосистему в її природному стані призводить до руйнування в ній генетико-еволюційних відношень. Так, у лучних та степових геосистемах еволюційно склалися відношення між рослинними угрупованнями та тваринами, за яких для нормального розвитку рослинності необхідне відчуження деякої частини її щорічної продукції травоїдними тваринами. Заповідання степових геосистем у режимі повного виключення стравлення та косіння призводить до досить- швидкої деградації рослинних угруповань, аж до випадання едифікаторних видів. Тому щоб підтримати заповідні степи в близьких до природних станах, необхідно ввести режим стравлення, близький до природно-еволюційного.
Врахування генетико-еволюційних відношень у геосистемах має значення для конструювання стійких і ефективних ландшафтів. Так, на цьому принципі має базуватися оптимальний підбір порід дерев для інтродукції, обгрунтування екологічно безпечних сівозмін тощо.