

§ 4.9. Межі між геосистемами
Коротка історична довідка. Тривалий час питання про межі в ландшафтознавстві зводилось до визначення їх місцеположення. Мабуть, першим, хто вважав межу специфічним і відносно самостійним предметом географічного аналізу, був В. П. Семенов— Тян-Шанський. У 1928 р. він звернув увагу на розпливчастість (континуальність) ландшафтних меж і запропонував формальний метод виділення перехідної смуги між природними районами та умовної лінійної межі на цій смузі. Проте належного відгуку в ландшафтознавців передвоєнного часу ці ідеї не знайшли.
Проблемою меж географи зацікавились у зв’язку з дискусією про дискретність-континуальність ПТК, яка розгорнулася в середині 50-х років. Сама ж концепція континууму та континуального (плавкого) характеру природних меж зародилася в фітоценології. Започаткував її Л. Г. Раменський. У 26-річному віці він зробив доповідь «Про принцип безперервності рослинного покриву» (1910), у якій обгрунтував положення про поступовість просторової зміни однієї рослинної асоціації іншою, умовність лінійних меж між ними. У цей же час аналогічні ідеї висловили в США Г. І'лізон, у Франції — Ф. Ленобль, в Італії — Г. Негрі. Ці погляди викликали запеклу критику численних прихильників дискретності рослинного покриву, які наполягали на чіткій відокремленості асоціацій у просторі, на об’єктивному існуванні лінійних меж між ними (В. М. Сукачов, Т. Фріз та ін.).
Затвердження концепції континууму у фітоценології пов’язано з іменами американських екологів Дж. Кертіса, Р. УІттекера, Р. Макінтоша, блискучими працями яких було доведено, що поступовість зміни природних умов у просторі та стохастичний зв’язок рослин з едафічними факторами зумовлюють поступовість переходу однієї асоціації в іншу, Згідно з твердженнями кконтинуалі стів» (до них належить більшість екологів), дискретні, тобто лінії, ні, межі в природі є, але вони зустрічаються рідко, з’являються внаслідок катастрофічних змін едафічних факторів і існують недовго, трансформуючись у перехідні смуги — екотони, або клінальні екосистеми. У ландшафтній екології та сучасному ландшафтознавстві переважають аналогічні погляди.
Типи ландшафтних меж. Одна геоснстема іншою може змінюватись двома шляхами: 1) стрибкоподібно вздовж певної лінії (лінійна, або дискретна межа); 2) займати деяку перехідну смугу, межі якої можна визначити (ландшафтний екотон).
Незалежно від ширини, ландшафтні межі можуть бути реальними («об’єктивними») та умовними. Місцеположення перших можна встановити однозначно. Прикладом таких меж є вододільні лінії у різкопочленованому ландшафті, тальвеги, екотони (перехідні смуги) на межі луків та лісу тощо. Умовні межі виділяють якдеякі лінії на реальній перехідній смузі, коли її необхідно умовно
зобразити у вигляді лінії (наприклад, на картах). Умовність їх лише в тому, що реальна ширина межі не береться до уваги, Межі, що розділяють між собою дві елементарні геосистемн (геотопи), можна назвати топічними ландшафтними межами, або межами 1-го рангу. Межі між геосистемами вищих рангів (мікро- геохор, ландшафтних смуг тощо) мають ранг, відповідний рангу контактуючих геосистем. Ранг межі пов’язаний з її контрастністю. Загалом, чим вищий ранг межі, тим вона контрастніша. Контрастність межі — її важлива характеристика, врахування якої дає змогу об’єктивізувати схеми ландшафтно-екологічного районування. Кількісно її можна оцінити за таким показником:
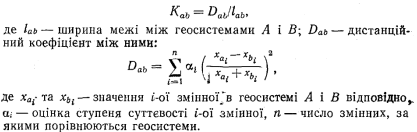
Морфологічна вираженість меж важлива при оцінці естетичної привабливості (атрактивності) ландшафту. За цією характеристикою межі можна розділити на морфологічно невиражені (наприклад, між геосистемами, що різняться між собою лише за грунтовими ознаками); слабковиражені (між рівнинними та полого- схиловими геосистемами, вододільні лінії на рівнинних); середньо- виражені (між увігнутою та опуклою частинами схилу); явно морфологічно виражені (бровки схилів, уступи та тилові шви терас). Морфологічна вираженість ландшафтних меж залежить від часу відособлення геосистем. Чим молодша територія, тим чіткіші межі (наприклад, у горах альпійської складчастості, у молодому ерозійному рельєфі). З часом внаслідок міжгеосистемних взаємодій морфологічна вираженість їх послаблюється.
За генезисом, тобто головним фактором, що зумовив появу межі, вони є літогенні, морфогенні, ледогенні, гідрогенні, фітогенні,, зоогенні, антропогенні. Проте переважна більшість їх комплексні (наприклад, морфолітогенні, педофітогенні тощо).
За функцією у ландшафтній територіальній структурі межі бувають контактними та бар’єрними (Каганський, 1982; Родоман^
. Вздовж контактних меж відбувається взаємодія двох сусідніх геосистем, взаємопроникнення їх властивостей, перенесення речовинно-енергетичних потоків. Такими є, зокрема, межі ландшафтних смуг на схилі, межі біоцентрів, ПГ-комплексш. Бар’єрна межа перешкоджає або повністю виключає взаємодію сусіднц геосистем. Яскравим прикладом їх є межі басейнів. Більшість меж по відношенню до різних типів горизонтальних міжгеосистемних зв’язків виконує і бар’єрну, і контактну функції. Д. Люрі (1985) запропонував називати їх мембранними.
Щодо напрямку горизонтальних динамічних потоків межі поділяються на дивергентні (межі розсіювання) — від них потоки- розходяться у різних напрямках (наприклад, вододільні лінії); конвергентні (межі-концентратори), вздовж яких різноспрямовані потоки зливаються (наприклад, лінії тальвегів); консеквентні — межі, які збігаються з лініями току (наприклад, лінії скатів схилів) ; градієнтні, вздовж яких помітно змінюється інтенсивність потоку (наприклад, лінії перегинів схилу, межі ландшафтних смуг).. Градієнтні межі поділяються на два підтипи — градієнтні імпульсні (вздовж них інтенсивність потоку зростає) та градієнтні гальмуючі (зменшується).
Ландшафтні екотони. Термін «екотон» увів в екологію Ф. Кле- ментс у 1928 р. Він розумів під ним перехідну смугу між двома досить контрастними екосистемами, через своєрідність якої їх не можна віднести ні до однієї із суміжних екосистем. Типовий приклад— смуга між лісом та степом. У ландшафтознавстві для таких смуг є й інші назви: «змінна ландшафтна одиниця» (Д. Л. Ар- манд, 1955); «буферна геоснстема» (В. Б. Сочава, 1978); «геотон»- (Н. Л. Беручашвілі, 1988).
Від клінальних геотопів ландшафтний екотон відрізняється тим, що в ньому формуються деякі риси, не властиві геосистемам які розділяються цим екотоном. У клінальних геосистемах таких рис немає, у їх межах властивості однієї геосистеми поступово переходять у властивості іншої. Екотон здебільшого вужчий за геосистеми, між якими він розташований, а клінальні геотопи можуть займати значні площі, нерідко більші від геосистем «ядерного, типу».
Виникнення ландшафтних екотонів зумовлене загальною закономірністю еволюції просторових систем — поступовим стиранням різких відмінностей (меж) у природі внаслідок все більшого розвитку міжгеосистемних взаємодій. Здебільшого лінійна межа з часом трансформується в перехідну смугу (чітко виражені бровки молодої ерозійної форми з часом руйнуються і стають все більш пологими поверхнями; лінійна межа між лісом і лукам» поступово згладжується взаємопроникненням лісових та лучних видів у суміжні біотопи і утвореная галявин). Подальший розвиток ландшафтного екотону призводить до формування в ньому деяких специфічних рис, не властивих жодній з контактуючих геосистем. З часом він перетворюється на клінальн.у, або «типову», геосисте- му з власними межами, можливо, також екотонного характеру.
Переважна більшість меж у ландшафті — екотони різної ширини. За периметром та розміром контактуючих геосистем естонські геоекологи Ю. Мандер та Ю. Ягомягі (1982) виділяють мік- роекотони (вони утворюються при контакті окремих парцел і геотопів до 40 м у діаметрі), мезоекотони (контакт ліс — луки, болото— ліс та ін.), макроекотони (виникають на межі великих лісових, болотних масивів, великих водойм тощо). Г. Вальтер (1976) виділяє також зоно-екотони (наприклад, зона лісостепу, субальпійський пояс), проте цими одиницями ландшафтна екологія здебільшого не займається.
Найкраще екотонні ефекти проявляються на контакті лісових геосистем з трав’яними (лучними, степовими, ріллею). Тому основні властивості ландшафтних екотонів розглянемо на цьому прикладі, грунтуючись на результатах досліджень Д. Л. Люрі (1986,
. Екотон між лісом та степом відіграє бар’єрну і контактну функції. Бар’єрна може реалізуватись трьома способами: 1) ско- тон як бар’єр-трансформатор впливає на перетинаючий його го- ризонтальний потік так, що характеристики потоку суттєво змінюються при досягненні суміжної геосистеми (наприклад, атмосферні потоки тепла і водяної парн в напрямку з лісу до степу в межах екотону нагріваються і висушуються, а в зворотному охолоджуються і зволожуються); 2) екотон як бар’єр-перешкода не дозволяє деяким потокам досягти суміжної геосистеми, при цьому матеріал, що переноситься потоком, акумулюється в межах екотону і далі включається в міжелементні зв’язки його вертикальної структури. Такими потоками, зокрема, є повітряне перенесення насіння, опаду, перенесення снігу хуртелицею, води поверхневого стоку тощо; 3) екотон як бар'єр-відштовхувач повертає горизонтальні потоки, які йдуть до нього від ядер суміжних геосистем (наприклад, зоогенні міграції типово лісових або типово степових видів).
Контактна функція ландшафтного екотону може бути реалізована у вигляді: простого контакту, коли горизонтальні потоки без ^перешкоди і видозміни перетинають екотон; активного контакту, коли в екотоні формуються нові потоки, невластиві ядрам типовості контактуючих геосистем (наприклад, атмосферні потоки на галявині—«бризові лісові вітри»); вторинного контакту, який проявляється в тому, що матеріал, накопичений в екотоні, починає мігрувати за його межі до суміжних геосистем.
Виділивши ландшафтний екотон між двома геосистемами, ви- .діляють і їх ядра типовості (екотон відтинає ці ядра). Визначати місцеположення ядер типовості потрібно при дослідженні ландшафтної динаміки, обгрунтування оптимального розташування .пунктів моніторингових спостережень та інших тестових ділянок ландшафтно-екологічних досліджень. Відносно цього ядра можна визначити ступінь типовості будь-якої точки геосистеми:
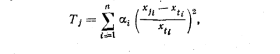


20, 25. Питання ландшафтного картографування докладно розглянуто в підручниках з ландшафтознавства (див., наприклад, Іса- ченко, 1991).
Графічні та матричні моделі. ЛТС будь-якого типу можна зобразити графом, вершинами якого є геосистеми, а ребрами — просторові відношення між ними.
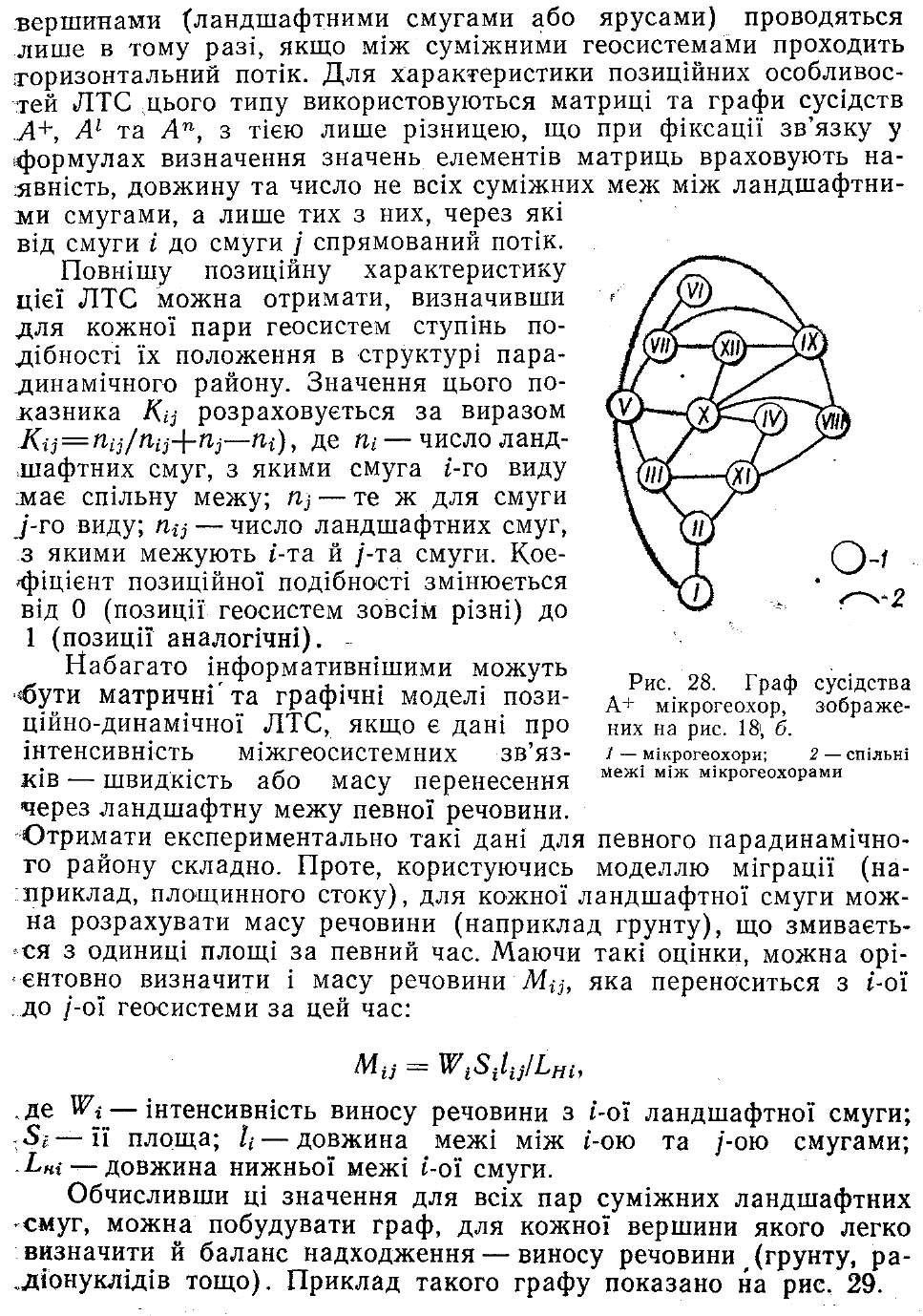
Для генетико-морфологічної ЛТС визначальними є відношення генетичної спільності між її геосистемами, Ця спільність тим більша, чим подібніші між собою геосистеми за комплексом генетично зумовлених ознак. Тому ступінь генетичної близькості двох геосистем можна визначити за допомогою певного дистанційного коефіцієнта. Обчисливши значення цього коефіцієнта для всіх можливих пар геосистем, отримаємо матрицю їх близькості. На її основі легко скласти граф подібності (рис. 27).
Важливі морфологічні особливості генетико-морфологічної ЛТС розкриваються через аналіз сусідства її геосистем. Цей аналіз зручно вести на основі матриць сусідств (синонім — матриця су-


Графи басейнової ЛТС. У гідрології та геоморфології традиційно прийнято зображати басейн річки за допомогою графу-де- рева, ребрами якого е водотоки, вершинами — точки їх злиття та витоку. За такими графами легко визначити порядок водотоку, число водотоків у межах будь-якого басейну чи його частини (правої або лівої), визначити деякі топологічні характеристики річкової сітки. Для ландшафтно-екологічного аналізу більш зручні графи, ребрами яких є не водотоки, а відношення підпорядкування басейнів, які показуються на графі як його вершини (див. рис. 2.3, ).
У графах біоцентрично-сітьової ЛТС вершинами є біоцентри,. ребрами — біокоридори та інтерактивні елементи. Граф може бути орієнтованим, якщо є вагомі підстави зважати на суттєву перевагу міграції видів лише, в певному напрямку. Ребрам можна також присвоїти чисельні значення, що оцінюють інтенсивність міграції вздовж біокоридору. Якщо ж емпіричних даних про інтенсивність міграції немає, її орієнтовно можна визначити за гравітаційною моделлю

У матричній формі біоцентрично-сітьову ЛТС можна показати у вигляді матриці суміжностей, елементами якої будуть 1 (коли між біоцентрами є біокоридор) та 0 (біокоридору немає), або значення коефіцієнту Сіj. Для оцінки ролі окремих біоцентрів у сітьовій структурі, важливій при визначенні природоохоронних пріоритетів, доцільно скласти матрицю доступності графу. Елементи aij такої матриці чисельно дорівнюють числу біокоридорів між біоцентрами і та / (тобто числу біокоридорів, які необхідно пройти з біоцентру і, щоб дістатися до біоцентру ). Графу біоцентрично- сітьової ЛТС, зображеному на рис. 26, б, відповідає матриця доступності, наведена в табл. 14. На її основі розраховуються важливі кількісні показники біоцентрично-сітьової ЛТС.
Кількісні показники. Для генетик о-м орфологічної ЛТС важливими є показники ступеня її однорідності — різноманітності. Цю особливість ландшафтної структури можна аналізувати в кількох аспектах і, відповідно, характеризувати різними показниками.
Доцільно розрізняти хорологічну різноманітність певного регіону, яка визначається кількістю контурів геосистем у ньому, та типологічну різноманітність, як функцію від кількості видів гео- систем у регіоні. Складність ЛТС — її характеристика, що поєднус




