

1avdin_v_v_matematicheskoe_modelirovanie_ekosistem
.pdfвелика роль тропических широт в глобальном балансе тепла и влаги и, следовательно, в общей циркуляции атмосферы.
Важным звеном в круговороте воды является использование водных ресурсов на нужды человека: население, промышленность сельское хозяйство. Наиболее ощутимые последствия антропогенного использования – загрязнение воды и безвозвратное водопотребление.
В последнее время возрастает использование на нужды человека океанских вод. Например, в Кувейте суммарная мощность опреснительных установок более
250 000 м3/сут.
Изменение осадков определяется в основном изменениями температуры. При изменении среднегодовой глобальной температуры атмосферы у поверхности Земли знак изменения среднегодовой температуры во всех зонах планеты будет одинаковым. Изменение режима осадков при этом происходит по-разному. Вблизи экватора при увеличении температуры количество осадков увеличивается, в районах средних широт – уменьшается. Кроме того, увеличение температуры приводит к увеличению нестабильности годового количества осадков в средних широтах. Эти факты имеют место при небольшом изменении температуры. При значительном же её повышении может возникнуть климат каменноугольного периода, при котором всюду было очень тепло и много осадков. Чтобы проанализировать вероятные ситуации, проведём предварительный анализ следующей модели.
Рассмотрим два важнейших эффекта, связанных с увеличением температуры: 1) увеличение испаряемости с поверхности суши и океана и как следствие увеличение общего количества влаги в атмосфере; 2) уменьшение градиента температур между экватором и полюсом. При этом, например, увеличение температуры у экватора может составить 0,5°С, а на полюсах – 1–2°С и более. Это означает, что меридиональный перенос влаги будет уменьшен. При таком подходе изменение осадков связано с двумя процессами: увеличением их суммарного количества и уменьшением скорости передвижения влаги к полюсам.
Пусть среднегодовая температура атмосферы у поверхности Земли Т определяет разность температур между экватором ТЭ и полюсом ТП ∆ТЭП = ТЭ – ТП. Рассматриваем только меридиональный перенос осадков от экватора к полюсам. Считаем, что испарение воды происходит только на экваторе с интенсивностью ЕЭ = ЕЭ(Т). Обозначим через МА = МА(ϕ, Т) мгновенное количество влаги в атмосфере, приходящейся на 1° географической широты ϕ. Скорость продвижения влаги от экватора к полюсам зависит только от ∆ТЭП и, следовательно от Т: Vϕ = g(N). Интенсивность выпадения осадков в данном месте
W(ϕ, T) = kMA,
где k – коэффициент пропорциональности, ϕ – географическая широта.
Считаем, что с увеличением средней глобальной температуры атмосферы испарение увеличивается, а скорость переноса влаги от экватора к полюсам уменьшается: dЕЭ/dТ > 0, dg/dT < 0. Будем также полагать, что выпавшие до 80° с. ш. и ю. ш. осадки попадают сразу обратно в океан, а оставшиеся осадки выпадают между полюсом и 80°, превращаются в лёд и снег, и попадают в океан в северном полушарии
41
в процессе таяния льдов, в южном – в виде айсбергов. Этим модель замыкается по количеству воды.
Пусть температура постоянна. В стационарном состоянии при движении по меридиану объём влаги, приходящийся на 1° широты имеет баланс
dM A |
= |
∂M A |
dϕ |
= |
∂M A g(t) = −W = −kM A , |
|
|
||||
dt |
∂ϕ dt |
|
dϕ |
||
откуда после интегрирования получим MA(ϕ) = MA(0) exp(–kϕ/g). Мгновенная масса осадков одного полушария
90 |
90 |
Wобщ = ∫ kM A (ϕ)dϕ = k ∫ M A (ϕ)dϕ = kMобщ, |
|
0 |
0 |
где Мобщ – общее количество влаги в атмосфере. В стационарном состоянии
dMобщ/dt = ЕЭ – Wобщ = ЕЭ – kMобщ = 0,
откуда Mобщ = ЕЭ/k. В то же время,
90 |
|
|
|
|
|
|
|
|
|
|
|
g |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Mобщ = ∫ M A (0)exp |
− kϕ dϕ = M A (0) |
. |
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
0 |
|
|
|
|
|
g |
|
|
|
|
k |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Таким образом, Mобщ |
= |
ЕЭ |
= M(0) |
g |
или |
M(0) |
= |
ЕЭ |
|
. Тогда |
||||||||||||||
|
|
g |
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
k |
|
k |
|
|
|
|
|
|
|||||||
E |
Э |
(Т) |
|
|
|
− kϕ |
|
|
|
|
|
E |
Э |
(Т) |
|
− kϕ |
||||||||
M A (ϕ,T) = |
|
|
|
exp |
|
, |
W(ϕ,T) = k |
|
|
|
exp |
. |
||||||||||||
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
g(T) |
|
|
g(T) |
|
|
|
|
|
|
g(T) |
g(T) |
||||||||||||
Отметим, |
|
|
что |
|
на |
экваторе |
|
(ϕ = 0) |
|
W = kEЭ/g и, значит |
||||||||||||||
∂W / ∂T = k(E′ g |
− E |
Э |
g′)/ g2 > 0, то есть при увеличении температуры осадки на эк- |
|||||||||||||||||||||
|
Э |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
ваторе всегда возрастают.
Рассмотрим стационарное распределение осадков при постоянных температурах. Направление изменения стационарного значения осадков в данном участке поверхности при изменении температуры определяются знаком производной
∂ |
W |
|
k |
|
|
|
k |
ϕ |
|
|
= |
e−kϕ/g E′Эg − g′EЭ 1 |
− |
|
. |
||||
∂T |
2 |
|
|
||||||
|
g |
|
|
|
g |
||||
Широта, на которой стационарное количество осадков не меняется (ϕ°), определяется из соотношения ∂W/∂T = 0, то есть ϕ°= (g / k)[1− E′Эg / EЭg′]. Но так как
согласно гипотезе E′Э > 0 и g′ > 0, то ϕ° > 0, то есть всегда существует место, где стационарная интенсивность осадков не изменится при варьировании температуры.
42
Заметим, что это место может быть и при ϕ > 80°. Легко показать, что при ϕ° < ϕ ∂W/ ∂T < 0, а при ϕ° > ϕ ∂W/∂T > 0, то есть с увеличением температуры между ϕ° и экватором осадки увеличиваются, а между ϕ° и полюсом – уменьшаются. Направление смещения точки ϕ° при изменении температуры определяется знаком ∂ϕ°/∂T.
Легко показать, что если g(T) и EЭ(T) – линейные функции температуры, то ∂ ϕ°/∂T < 0. Это означает, что при увеличении температуры стационарное значение, при котором количество осадков не меняется, будет смещаться к экватору.
Рассмотрим, каким будет изменение режима осадков при увеличении температуры относительно некоторого «современного» значения. Изучим предельные изменения осадков. Если lim g(t) = 0 , то распределение осадков будет стремиться к
t→∞
положению, когда все осадки выпадают в месте испарения воды. Если при повышении температуры, начиная с некоторого Т = Т*, скорость переноса осадков перестаёт уменьшаться, то есть g(T) = B = const при Т > Т*, то получим «предельное» распределение:
W(ϕ,T) = |
kEЭ(T) |
|
− |
kϕ |
= f(ϕ,T). |
|
|
exp |
|
|
|||
B |
|
|||||
|
|
|
B |
|
||
При этом осадки с температурой увеличиваются при любом значении ϕ, то есть эта ситуация соответствует условиям каменноугольного периода.
Принебольших изменениях температуры ипри∂g/∂T < 0 возможны дваслучая. 1. Современное значение 0° < ϕ° < 80°. Тогда при увеличении температуры осадки между экватором и ϕ° увеличатся, а между ϕ° и полюсом – уменьшатся. При этом, как было показано, точка неизменного их количества будет смещаться к эква-
тору.
2. Современное значение ϕ° > 80°. Тогда при небольшом увеличении температуры осадки почти всюду, от 0 до 80° увеличатся. С дальнейшим возрастанием температуры точка сместится до значения 0° < ϕ° < 80°, и изменение осадков будет происходить в соответствии с первым случаем.
11. МОДЕЛИРОВАНИЕ ОКЕАНИЧЕСКОЙ БИОТЫ
Рассмотрим структуру экосистем океана. На долю океана приходится значительная часть процесса фотосинтеза, и по имеющимся оценкам общее количество органического вещества, продуцируемого в океане, составляет около 30 % органического вещества, продуцируемого растениями суши. Сопоставление продуктивностей экосистем океана и суши в настоящее время оказывается не в пользу первых. Океан занимает 71 % поверхности планеты, а даёт всего лишь около 1 % от общего количества потребляемых человечеством пищевых продуктов. Объясняется это интенсивным культурным хозяйством, существующим на суше и практически отсутствующем в океане, кроме того, характером исторического развития. Это вызвано значительными отличиями структуры океанских экосистем, для которых, в отличие от наземных, типично большое количество трофических уровней, пространственная неустойчивость и большая уязвимость.
43
Жизнь в океане распределена весьма неравномерно. Наиболее богатые жизнью океанские воды находятся в мелких местах – на отмелях, атоллах, континентальных шельфах и др. Это однозначно определяется характером цикла жизни в океане, когда лимитирующими факторами являются в основном солнечный свет и биогенные элементы, а её первоисточником служит продукция одноклеточных – фитопланктона.
Можно предположить, как и ранее, что небольшие изменения глобальной температуры атмосферы будут сопровождаться изменением температуры на каждой широте в том же направлении. Тогда при увеличении температуры прогревание верхнего слоя океанских вод, зависящее от температуры воздуха, изменится таким образом, что зоны наибольшего сезонного перемешивания (обуславливающие приток питательных веществ и, следовательно, высокую плотность жизни) сместятся к высоким, ранее более холодным широтам. Уменьшение глобальной температуры сместит зоны интенсивной океанской жизни в сторону экватора. Следовательно, приближённо можно считать, что изменение температуры атмосферы может привести к смещению зон жизни в океане, но не к изменению их площади.
Эффективность управления продуктивностью океана существенно зависит от пространственной и трофической структуры населяющих его сообществ, которые определяются комплексом природных и антропогенных факторов. При построении модели необходимо учитывать тот факт, что вдоль всех континентов тянутся «прилежащие зоны» – районы открытого моря, прилегающего к территориальным водам, в пределах которых государство вправе осуществлять контроль над использованием ресурсов океана. То есть вся территория шельфа, где наиболее высока продуктивность океана, находится под контролем разных государств.
В настоящее время в океане преобладают экосистемы с естественными структурами, выработанными в процессе эволюции. Они существенно различаются по видовому составу и форме трофического графа. Однако все они являются открытыми системами, диссипативными структурами, на вход которых поступает энергия солнечной радиации, а на выходе – продукция высших трофических уровней сообщества. Эта продукция зависит от количества входящей в него энергии и от эффективности её использования в пищевой сети сообщества. При этом от данных факторов зависит устойчивость экосистемы и её выживаемость. Продукция экосистемы в общем виде определяется как P = kE, где k – передаточный коэффициент, Е – энергия солнечной радиации. k – безразмерен, следовательно продукция оценивается в энергетических единицах. Наиболее устойчивыми являются системы, у которых низок передаточный коэффициент k. Устойчивость и продуктивность для океанских экосистем находятся в обратной зависимости.
Построение океанского блока моделей требует детального математического описания совокупности происходящих в экосистемах океана процессов энергообмена между трофическими уровнями и взаимосвязей биотических, абиотических и гидрологических факторов. Схематически эти связи представлены на рис. 9.
Будем считать, что в океане единственным источником энергии и вещества для всех форм жизни служит первичная продукция фитопланктона. Из вне поступает только энергия солнечной радиации Е. Первичная продукция в этом случае лимитируется освещённостью и концентрацией биогенных элементов. Рассмотрим отдельные зависимости.
44

Антропогенные факторы |
|
|
Гидро- |
Абио- |
|
логические |
тические |
|
условия |
условия |
Живые |
|
|
компоненты |
|
|
кормовой |
|
|
базы |
|
Нектон |
|
Мёртвое органическое вещество |
||
Рис. 9. Взаимосвязь между живыми и косными компонентами |
||
океанической экосистемы |
|
|
Зависимость освещённости от глубины. Для данной зависимости существует большое количество экспериментального материала. Аппроксимация этой зависимости математической функцией позволяет записать следующее эмпирическое уравнение:
|
n |
z |
|
, |
EZ = E(t, x, y, z) = E(t, x, y,0)exp |
− αz − ∑λi ∫ Bi (t, x, y, u)du |
|||
|
i=1 |
0 |
|
|
где x, y, z – координаты, u – некоторая функция освещённости, α – коэффициент поглощения света чистой водой, Bi – биомасса i-го вида, λi – показатель изменения освещённости за счёт затенения биомассой В. На рис. 10 показаны теоретические (штриховые) и экспериментальные (сплошные) кривые изменения освещённости с глубиной. 1– средняя для тропиков, 2 – южные тропики, 3 – в Ионическом море, 4 – в западной части Тихого океана.
45
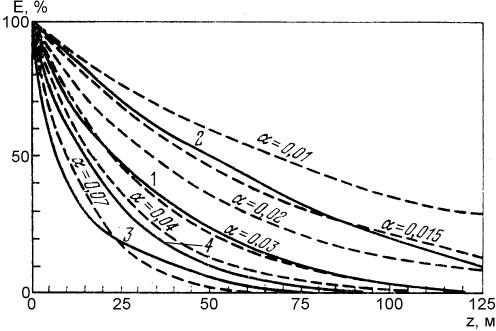
Рис. 10. Зависимость освещённости от глубины
Фотосинтез. Скорость фотосинтеза как функция освещённости имеет один максимум при некотором оптимальном значении Е = Е*, убывая при отклонении освещённости от этого критического значения (рис. 11). Это обстоятельство приводит к тому, что интенсивность фотосинтеза в океанских глубинах, в отличие от освещённости, меняется по закону, описываемому кривой с одним максимумом (рис. 12), соответствующем глубине zmax, определяемой условием E(t, x, y, zmax) = E*(t, x, y). Сплошная линия – экспериментальные данные, пунктир – расчёт. В различных географических широтах положение этого максимума может значительно изменяться.
В тропических зонах максимум фотосинтеза в разные сезоны года располагается на глубинах 10–30 м, в редких случаях (в открытых районах с повышенной прозрачностью воды) – на глубине ниже 30 м.
Согласно имеющимся экспериментальным данным, максимальная скорость фотосинтеза в сентябре – октябре в экваториальных районах Атлантического океана наблюдалась при интенсивности солнечной радиации 65–85 кал/см2 в день. Начиная с глубины, где энергия солнечной радиации составляла 20–25 кал/см2, фотосинтез уменьшался пропорционально общей энергии радиации. Уменьшение скорости фотосинтеза наблюдалось при энергии радиации выше 100 кал/см2, когда происходило явное угнетения фитопланктона светом.
Толщина фотосинтезирующего слоя изменяется примерно от 50 м у побережья до 100 м в открытом океане. При этом положение слоя максимального фотосинтеза колеблется от 0 до 10 м в прибрежной зоне и от 5 до 30 м – вдали от берегов. Соответственно интенсивность фотосинтеза в среднем для глубин 0,5 и 100 м характеризуется значениями 200 и 0,2 мгС/(м3 сут) вблизи берегов и значениями 6,2 и 0,3 мгС/(м3 сут) в открытом океане.
Фитопланктон разных морей и широт обладает различными световыми характеристиками и степенью адаптации к использованию света. Региональные различия учитываются в модели путём изменения соответствующих коэффициентов в уравнении фотосинтеза.
46

Рис. 11. Зависимость скорости фотосин- |
Рис. 12. Зависимость скорости фотосин- |
теза от освещённости |
теза о глубины |
Скорость фотосинтеза на глубине z зависит от освещённости, температуры, концентрации биогенных элементов (В12), биомассы фитопланктона (В2) и других факторов. Для выражения этой зависимости используются различные уравнения, которые в той или иной форме отражают лимитирующую роль факторов. Для выражения этой зависимости используются различные уравнения, которые в той или иной форме отражают лимитирующую роль факторов. При этом, очевидно, интенсивность фотосинтеза, вычисленная по таким зависимостям, должна стремиться к нулю при уменьшении освещённости, концентрации биогенных элементов и фитопланктона. Увеличение концентрации биогенных элементов должно приводить к эффекту насыщения. С учётом сказанного скорость фотосинтеза будет выражаться как R2 = min( ρ2B2, R22), где R22 определяется как
R |
|
= k |
|
e−b2 / B2 |
[1− exp(− k |
B |
)]Eexp 1− |
E |
e−d2θZ |
1+ α2T |
∆T |
|
, |
|
22 |
2,0 |
|
|
|||||||||||
|
|
|
|
2,12 12 |
|
|
|
|
100 |
|
|
|||
|
|
|
|
|
|
|
|
E* |
|
|
|
|
||
где ρ2 – максимальный П/Б-коэффициент фитопланктона (отношение продуктивности к биомассе); d2 и α2Т – показатели влияния на фотосинтез загрязнения θZ и изменений глобальной температуры ∆Т соответственно; k2,12 – индекс чувствительности фотосинтеза к концентрации биогенных элементов; k2,0 – коэффициент пропорциональности, который можно выразить через ассимиляционное число Аmax определяется из выражения k2,0 = k0Amax/E*, где, в частности, можно принять:
5,9E* в области максимального фотосинтеза,
Amax =
2,7E* для" теневого" фитопланктона.
47

Скорость фотосинтеза также существенно зависит от температуры среды и возрастной структуры фитопланктона. Известно, что удельная интенсивность фотосинтеза фитопланктона с изменением температуры от низкой к более высокой сначала возрастает, достигая в некотором диапазоне оптимальных для В2 температур наивысшего значения, а затем с дальнейшим повышением температуры начинает падать.
Зависимость интенсивности фотосинтеза от возраста определяется комплексом многочисленных факторов; для простоты её можно считать убывающей. Например, для тропического фитопланктона ρ2 может меняться от 5 до 1, убывая до единицы на 15–17 сутки существования.
Влияние концентрации пищи на скорость роста потребляющего её компо-
нента. Рост консумента за счёт съеденной пищи определяется его биомассой и величиной суточного рациона. Количество пищи в случае её недостатка может служить лимитирующим фактором, а при её избытке, из-за физиологической ограниченности максимального суточного рациона не может привести к увеличению скорости роста больше некоторого порогового значения. Поэтому функциональную зависимость рациона R от биомассы пищи P можно аппроксимировать следующим выражением:
R = Rmax[1–exp(–kpP)], |
(40) |
где Rmax – максимальный рацион при достаточном количестве пищи, соответствующий полному удовлетворению пищевых потребностей.
Такая запись функции R хорошо описывает реальную зависимость для рыб. Значение kp в формуле (40) определяется тем, до какой величины P должна возрасти концентрация пищи, чтобы рацион составил некоторую заданную долю ν, чаще всего 0,99 от максимального. Тогда
kp = − P1 ln(1− ν) .
Пусть пищевой спектр j-го компонента экосистемы составляет множество Sj и пусть минимальная потребляемая биомасса i-й пищи составляет величину Pi, min. Тогда согласно (40) имеем
|
|
|
|
|
|
|
|
|
|
|
|
|
|
, |
(41) |
|
|
|
|||||
R j = R j,max 1− exp |
− ∑k ji Pi |
||||||
|
|
i Sj |
|
|
|
|
|
|
|
|
|
|
|||
где Pi = max{Pi − Pi,min ; 0}. Коэффициенты kij в (41) могут вообще быть различными для разных видов пищи спектра Sj и тем самым через них можно отражать эффективность i-й пищи. В целом сумма Vi = ∑k ji Pi представляет собой всю эффектив-
i Sj
ную пищу j-го компонента.
Уравнение баланса веществ и энергии. В любой экосистеме океана поток ве-
щества и энергии проходит через трофический граф и подчиняется следующему закону:
48

∂Bi = Ri − Hi − Ti − Mi − ∑G ji R j , |
(42) |
|
∂t |
jΓi |
|
где Гi – множество трофической подчинённости i-го компонента системы; Ri, Hi, Ti, Mi – соответственно величины неусвоенной пищи, затрат на энергетический обмен и отмирание (переход живой биомассы в мёртвое органическое вещество), Gji – доля i- го компонента в пищевом спектре j-го компонента, Вi – биомасса i-го компонента.
Полагая, что изъятие биомассы жертв производится хищником с интенсивностями, пропорциональными их биомассам, для коэффициентов Gji можно предложить следующее выражение:
G ji = k ji Bi ∑k jm Bm , m Sj
где Bi – значение биомассы i-го компонента, при которой рацион составит некоторую долю ν = 0,99 от максимального, Bm – значение усвоенной биомассы j-м ком-
понентом.
Степень усвояемости пищи может быть оценена по разности между содержанием органического вещества в потреблённой пище и в фекалиях:
P = |
Pr − Ph |
|
100% , |
(43) |
||
|
|
|||||
u |
(1 |
− P |
)P |
|
|
|
|
|
h |
|
r |
|
|
где Pr и Ph –доли органического вещества соответственно в пище и фекалиях. Расчёт величины Нi в (42) можно произвести на основе коэффициента ис-
пользования энергии усвоенной пищи k2 = П/(П+Т), где П – прирост биомассы консумента (в энергетических единицах), Т – затраты на обмен.
Согласно имеющимся экспериментальным данным, усвояемость пищи зоопланктоном постоянна в пределах её концентрации 20–30 мг/л, а в остальных случаях зависит от концентрации. Коэффициент использования потреблённой пищи на рост (k1) в среднем составляет 25,4 %, а коэффициент k2 – около 40 %. Усвояемость u–1 – доля биомассы жертвы, использованная хищником для увеличения собственной биомассы. Значение u–1 составляет около 72,4 %. Все эти величины зависят также от структуры рациона.
Таким образом, Rmax можно определить как
R max = |
1 |
(Пmax + T) = |
T |
1 |
. |
(44) |
|
h |
h 1− k2, max |
||||||
|
|
|
|
||||
Сравнивая (43) и (44), видим, что Pu /100 = 1–h.
Функция Ti в (42) отражает величину потерь энергии гидробионтами за счёт дыхания. Интенсивность дыхания водных организмов неодинакова у представителей разных трофических уровней, меняется с возрастом и зависит от параметров окружающей среды. Для целей моделирования можно использовать общеизвестную зависимость величины газообмена от массы тела:
49
Ti = τi wωi ,
где τi – удельная интенсивность дыхания, w – масса гидробионта, ωi – показатель зависимости интенсивности газообмена от массы. ωi = 0,7–0,75 для пойкилотермных и ωi = 0,95 для мелких многоклеточных.
Из внешних условий наибольшее влияние на потребление кислорода гидробионтами оказывают температурные и респираторные. Согласно многочисленным исследованиям интенсивность дыхания гидробионтов с повышением температуры до некоторого предела возрастает и может быть приблизительно описана «нормальной кривой» Крога. При этом для каждого вида существуют два граничных температурных порога, в пределах которых величина газообмена не зависит от температуры. Этот интервал называется «температурной зоной адаптации». Для планктонных животных эта зона является довольно широкой за счёт существования у них компенсаторных механизмов.
С учётом сказанного в модели используется следующая аппроксимация для описания зависимости интенсивности дыхания от температуры:
* |
при t |
0 |
0 |
0 |
], |
|
|
|
τi |
|
[t1,i ;t2,i |
|
|
|
|||
τi (t0 ) = |
|
|
|
0 |
00 |
|
[t10,i ;t2,i0 ], |
|
τ*i |
[ai exp(−bi t0 )](t |
−ti )/mi при t |
0 |
|||||
где t0 – локальная температура воды, t10,i |
|
и |
t2,i0 – нижняя и верхняя границы темпера- |
|||||
турной зоны адаптации, |
ti00 |
= 12 (t10,i + t2,i0 |
) |
, ai, bi, mi, – постоянные величины, выби- |
||||
раемые из условий наилучшей аппроксимации экспериментальных данных, τ*i – ста-
ционарные удельные интенсивности дыхания.
Величина τi также зависит от концентрации растворённого в воде кислорода (В14). Эта зависимость обычно такова, что с падением концентрации кислорода ниже определённых значений интенсивность газообмена гидробионтов падает. Поэтому для её описания воспользуемся выражением
τ |
i |
(t0 ) при B |
> B* |
, |
|
|
|
||
|
|
14 |
|
14,i |
|
|
|
|
|
τi (B14 ) = |
|
(t0 )B |
/ B* |
|
при 0 |
≤ B |
≤ B* |
, |
|
τ |
i |
|
|||||||
|
14 |
14,i |
|
|
14 |
14,i |
|
||
где знак * означает стационарное состояние.
У фитопланктона потери на дыхание характеризуются снижением усвоения углерода при фотосинтезе. Для Перуанского течения эти потери в среднем довольно высоки – до 74,6 % от поглощённого углерода. В количественном отношении это
1,45 гС/(м2 сут).
Показатель смертности Mi в формуле (42) отражает изменение скорости прироста биомассы Bi за счёт перехода живой биомассы в детрит. Mi определяется как
M |
i |
= (µ |
i |
+ µ′B |
)B |
(m′ + m′′/ B |
)θti0 −ti0* , |
(43) |
|
|
|
i i |
i |
i |
i 14 |
|
|
||
50