

Заняття №1. Будова клітини. Класифікація та будова хромосом.
Мета заняття: ознайомити студентів з основними положеннями клітинної теорії, особливості збереження та передачі спадкової інформації на клітинному рівні, особливостями будови тваринної клітини, будовою та функціями її структурних одиниць.
Теоретичні положення.
На сучасному етапі розвитку цитології клітинна теорія включає такі положення:
- клітина - елементарна одиниця будови і розвитку всіх живих організмів;
- клітини всіх одноклітинних і багатоклітинних організмів подібні за походженням (гомологічні), будовою, хімічним складом, основними проявами життєдіяльності;
- кожна нова клітина утворюється виключно внаслідок розмноження материнської шляхом поділу;
- у багатоклітинних організмів, які розвиваються з однієї клітини - зиготи, спори тощо, - різні типи клітин формуються завдяки їхній спеціалізації впродовж індивідуального розвитку (онтогенезу) особини та утворюють тканини;
- із тканин складаються органи, які тісно пов'язані між собою й підпорядковані нервово-гуморальній та імунній системам регуляції.
Залежно від рівня клітинної організації розрізняють два типи організмів — прокаріоти і еукаріоти.
Прокаріоти. У прокаріотів, до яких належать бактерії і синьозелені водорості, клітини невеликих розмірів (0,5-3 мкм). Ці клітини позбавлені ядерної мембрани і не містять чітко оформлених, обмежених мембраною органел. У прокаріотичних клітинах відсутнє ядро і хромосоми. Генетичний матеріал представлений однією довгою кільцевою молекулою ДНК. Гістонових білків не виявлено, у них відсутня нуклеосомна організація хроматину. Кільцева молекула ДНК упакована в клітині у вигляді петель.
Прокаріотичні клітини, оточені клітинною стінкою, що складається головним чином із вуглеводів і амінокислот. Плазматична або клітинна мембрана часто утворює випинання в цитоплазму, які називаються мезосомами. Вони виконують функцію утворення АТФ - багатої на енергію сполуки. У прокаріотів рух цитоплазми й амебоїдний рух відсутні; переміщуються вони найчастіше за допомогою джгутиків, організованих значно простіше від джгутиків еукаріотів. Прокаріоти поширені практично всюди.
Вони відрізняються величезною різноманітністю, швидким ростом, коротким часом генерації. Прокаріотична клітина може зазнавати поділу через кожних 20 хв. і таким чином утворювати за 10 год до 5 млрд. клітин.
Деякі представники прокаріотів:
• Мікоплазми - найпростіші з клітин, діаметром близько 0,1-0,3 мкм. Вони є внутрішньоклітинними паразитами.
• Рикетсії - збудники епідемічного висипного тифу. Середні розміри їх 0,3-0,6 мкм.
• Коки - невеликі округлі бактерії (стафілококи, гонококи та ін.) Діаметр їх 0,8-10 мкм. • Бацили - паличкоподібні бактерії (кишкова паличка, сальмонела та ін.)
• Вібріони - за формою нагадують кому, завдовжки 1,5-3,0 мкм, завтовшки 0,3 мкм (холерний вібріон).
Основні відмінності між прокаріотами і еукаріотами
Параметри |
Прокаріоти |
Еукаріоти |
Розмір клітин |
Діаметр від 1 до 10 мкм, у середньому складає 0,5-2,0 мкм |
Діаметр від 8 до 100 мкм, у середньому 40-60 мкм |
Форма |
Одноклітинні або нитчасті |
Одноклітинні, нитчасті або багатоклітинні з диференціацією |
Генетичний матеріал |
Кільцева ДНК знаходиться в цитоплазмі, не зв'язана з білками, нічим не відділена від цитоплазми. Немає ядра та ядерця. Хромосома кільцева |
Довгі, лінійні молекули ДНК, зв'язані з білками. Ядро відділене від цитоплазми оболонкою, усередині ядра знаходиться ядерце. Хромосоми всередині ядра, оточені ядерною мембраною |
Синтез білків |
Рибосоми дрібні, чутливі до антибіотиків. Ендоплазматичного ретикулуму немає |
Рибосоми більші, можуть бути прикріплені до ендоплазматичного ретикулуму |
Органели |
Органел мало або вони відсутні. Жодна з них не має мембранної оболонки. Внутрішні мембрани зустрічаються рідко; на них перебігають процеси дихання або фотосинтезу |
Органел багато. Деякі з них оточені подвійною мембраною, наприклад, ядро, мітохондрії, хлоропласти. Багато органел, обмежених одинарною мембраною, наприклад, комплекс Гольджі, лізосоми, вакуолі, мікротільця, ендоплазматичний ретикулум тощо |
Клітинні стінки |
Жорсткі, містять полісахариди й амінокислоти. Основний компонент, що їх зміцнює, - муреїн |
У рослин і грибів клітинні стінки жорсткі і містять полісахариди. Основний компонент клітинної стінки, що зміцнює її, у рослин - целюлоза, у грибів - хітин. Клітини тварин не мають стінок |
Цитоплазма |
Немає цитоскелета, руху цитоплазми, ендо- й екзоцитозу |
Добре розвинений цитоскелет, рух цитоплазми, ендо- й екзоцитоз |
Джгутики |
Прості, мікротрубочки відсутні. Знаходяться поза клітиною (не оточені плазматичною мембраною). Діаметр 20 нм |
Складні, із розташуванням мікротрубочок типу 9-2. Розміщуються всередині клітини (оточені плазматичною мембраною). Діаметр 200 нм |
Дихання клітин |
У бактерій дихання анаеробне або аеробне, відбувається в мезосомах; у синьо-зелених водоростей – на цитоплазматичних мембранах |
Аеробне дихання відбувається в мітохондріях |
Поділ клітин |
Простий поділ клітин, немає мітозу |
Мітоз (або мейоз) |
Фотосинтез |
Хлоропластів немає. Відбувається на мембранах, що не мають специфічної упаковки |
Хлоропласти є в рослинних клітинах, що містять спеціальні мембрани, які упаковані в ламели або грани |
Вакуолі |
Відсутні |
Присутні |
Капсула |
Може бути |
Відсутня |
Еукаріоти. Еукаріоти - організми, клітини яких мають ядро (від грец. - κάρυον), оточене мембранною оболонкою. До еукаріотів належать найпростіші, гриби, рослини і тварини. Генетичний матеріал зосереджений переважно у хромосомах, що мають складну будову й утворені нитками ДНК і гістоновими білковими молекулами. Поділ клітин - мітотичний. У цитоплазмі розрізняють багато характерних органел: центріолі, мітохондрії, пластиди та інші. Серед еукаріот існують як одноклітинні, так і багатоклітинні організми, яким властивий складний принцип структурної організації. Форми клітин можуть бути різноманітними, розміри коливаються в межах - від 5 до 100 мкм. Клітини мають подібний хімічний склад і обмін речовин. Вони розподілені системою мембран на компартменти. Усі клітини мають єдину систему збереження та реалізації спадкової інформації.
Особливості збереження та передачі спадкової інформації на клітинному рівні. Певна організація живого зв'язана зі збереженням і використанням потоку інформації для підтримки структурно-функціональної організації клітини та її тривалого стабільного існування як системи. Спадкова інформація зберігається в молекулах ДНК у вигляді генетичного коду – послідовності триплетів нуклеотидів. Інформація переписується з ДНК на молекули РНК, що забезпечують синтез необхідних структурних білків і ферментів. Утворені білки причетні до появи певних властивостей клітини. Іншими словами, потік інформації в клітині спрямований від ДНК до ознаки:
ДНК => РНК => білок => ознака
Ще один інформаційний потік направлений від ДНК однієї клітини до ДНК дочірньої клітини. Цей потік пов'язаний із процесом розмноження. Він реалізується реплікацією молекул ДНК материнської клітини, утворенням хромосом, процесом рівномірного розподілу спадкового матеріалу між дочірніми клітинами (мітозом):
ДНК => 2 дочірні ДНК =>=> дві дочірні клітини
Цей потік інформації забезпечує відтворення і тривале існування популяцій клітин.
Еукаріоти — рослини, тваринні, гриби і інші організми, що володіють складно організованим ядром. Основними компонентами еукаріотічних клітин є клітинна оболонка, ядро з ядерцем і цитоплазма (рис.1).
Основними компонентами клітин є біомембрани (7), цитоплазма(11) і ядро (2). Організація і функціонування всіх компонентів клітини пов'язані в першу чергу з біологічними мембранами.
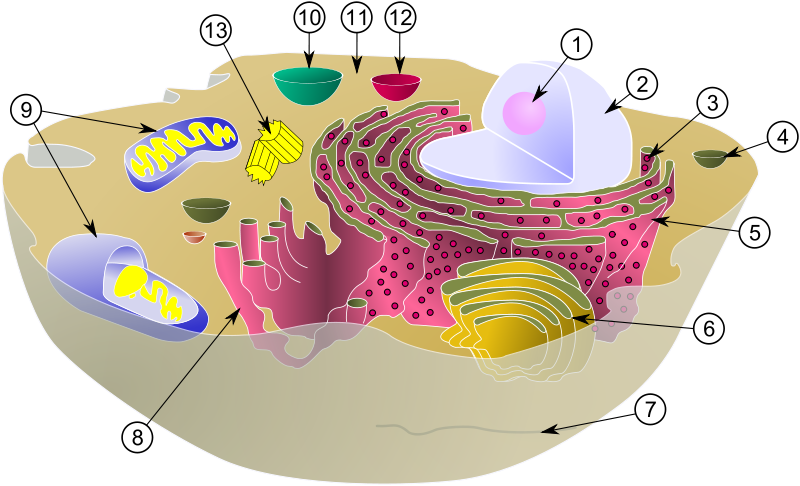
Рис.1. Будова клітини.
Сучасна цитологія розглядає біомембрани як один з основних компонентів клітинної організації, як основу структури і функцій всіх органів і тканин.
Більшість клітинних органел мають у своїй основі мембранні структури. Вони характерні для ендоплазматичної сітки, пластинчастого комплексу Гольджі, оболонок і крист мітохондрій, лізосом, вакуоль, пластид, ядерної оболонки і зовнішньої клітинної мембрани.
Мембрани - високовпорядковані, складні молекулярні системи, відповідальні за основні процеси життєдіяльності клітин. Наприклад, мембрани поділяють вміст клітини на відсіки (колтартменти), завдяки чому в клітині одночасно можуть перебігати різні, навіть антагоністичні, процеси; регулюють метаболічні потоки; підтримують різницю концентрацій речовин (іонів, метаболітів) шляхом переміщення; створюють різницю електричних потенціалів; беруть участь у процесах синтезу і каталізу та ін. Крім того, мембрани є основою для точного розміщення ферментів, а тому зумовлюють впорядкованість обмінних реакцій. Так, в ендоплазматичній сітці відбувається синтез білків, жирних кислот і фосфоліпідів.
У мітохондріях здійснюється цикл Кребса, окисне фосфорилування, окиснювання жирних кислот. Існує кілька типів мембран, які відрізняються за будовою, ферментативними властивостями білків, містять різні ліпіди. Так, мембрани мітохондрій тонкі (близько 5 нм) і мають глобулярну структуру білків і специфічний набір фосфоліпідів. Мембрани комплексу Гольджі досить товсті (6-9 нм), містять інші білки і ліпідні молекули. У цитоплазматичних мембранах знаходяться молекули-рецептори до біологічно активних сполук, наприклад, гормонів. Більшість захворювань людини і тварин пов'язані з порушеннями будови і функції мембран.
Клітина мембрана. Відповідно до рідинно-мозаїчної моделі будови, клітинні мембрани - це напівпроникний ліпідний бішар із вбудованими в нього білками (рис. 2).
Мембрани різних органел мають неоднаковий ліпідний і білковий склад, що забезпечує їх функції. Кожний різновид мембран містить близько 50 % білків. Мембрани мають також значний відсоток вуглеводів. Наприклад, мембрана еритроцитів складається з 40 % ліпідів, 52 % білків і 8 % вуглеводів. Білки не утворюють шари, а розташовані нерівномірно у вигляді мозаїки з глобул; при цьому одні з них знаходяться тільки на поверхні, інші занурені в ліпідну фазу частково або повністю, іноді пронизують її наскрізь.
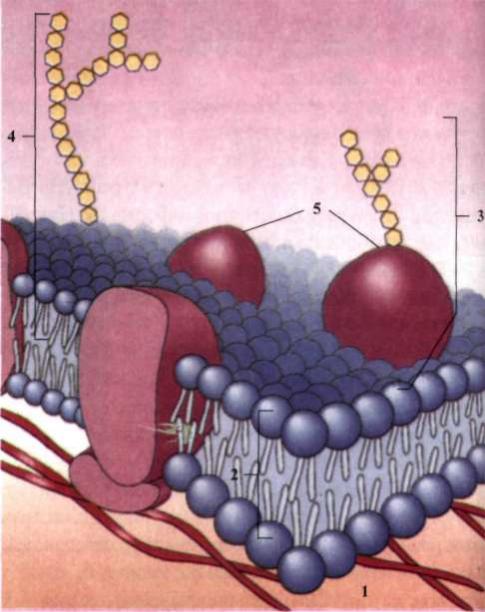
Рис. 2. Будова клітинної мембрани: 1 - цитоплазма; 2 - ліпідний бішар; З - глікопротеїн; 4 - гліколіпід; 5 - протеїни.
Ліпідний бішар являє собою рідину, в котрій окремі молекули ліпідів здатні дифундувати в межах свого моношару, але можуть іноді переміщатися з одного шару в другий. Цитоплазматична мембрана зовні вкриває клітину і є важливою ланкою в системі біомембран, необхідною умовою існування будь-якої клітини. Її поява була однією з умов виникнення життя.
До складу цитоплазматичних мембран, крім ліпідів і білків, входять також молекули гліколіпідів і глікопротеїдів із розгалуженими вуглеводними ланцюгами. Ці розгалужені ланцюги на поверхні клітини переплітаються один з одним, створюють ніби каркас із вплетеними в нього молекулами білків (глікокалікс), що складається з полісахаридів, ковалентно зв'язаних із глікопротеїдами і гліколіпідами плазмолеми. Функції глікокаліксу: а) міжклітинне розпізнавання; б) міжклітинна взаємодія; в) пристінкове травлення.
Функції біологічних мембран у значній мірі визначають властивості і фізіологію клітини.
Транспорт речовин через мембрану. Ліпідний бішар практично непроникний для більшості полярних водорозчинних молекул, оскільки внутрішня частина його гідрофобна. Завдяки такому бар'єру запобігається втрата водорозчинного вмісту клітини. Різні речовини мають неоднакову здатність проникати через цей бар'єр. Великі макромолекули (білки, жири) та їх агрегати не можуть проникати крізь мембрану. Для перенесення існує "макромеханізм" – захоплення клітиною і доставка в певному напрямку (ендоци-тоз і екзоцитоз). Невеликі молекули переносяться за допомогою спеціальних молекулярних механізмів через мембрану: шляхом пасивного й активного транспорту.
Транспорт невеликих молекул. Є два типи транспорту молекул через мембрану: пасивний і активний.
Пасивний транспорт - переміщення невеликих полярних (СО2, Н20) і неполярних (02, N ) молекул за градієнтом концентрації або електрохімічним градієнтом без витрати енергії. Існують різні форми пасивного транспорту:
1. Проста дифузія
2. Полегшена дифузія
3. Осмос
Активний транспорт - перенесення молекул через мембрану за допомогою спеціальних білків проти концентраційного або електрохімічного градієнта з використанням енергії АТФ. Білки-переносники є одночасно ферментами і називаються АТФ-азами.
Транспорт агрештш великих молекул. Ендоцитоз - складний активний процес поглинання клітиною великих молекул, часток, мікроорганізмів. Різновиди: піноцитоз, фагоцитоз, опосередкований рецепторами ендоцитоз. Піноцитоз - поглинання рідини та розчинених речовин з утворенням специфічних мембранних пухирців.
Цитоплазма складає основну масу клітини – це весь її внутрішній вміст, за винятком ядра. Містить 75-85 % води, 15-25 % білків і багато інших речовин, але в менших кількостях. При вивченні клітини за допомогою світлового мікроскопа цитоплазма є гомогенною, безбарвною, прозорою, в'язкою рідиною.
Проте електронний мікроскоп дозволив виявити складну багатокомпонентну, поліфункціональну, високовпорядковану структуру цитоплазми. Цитоплазма складається із цитозолю (цитоплазматичний матрикс), внутрішньоклітинних органел і включень.
Цитозоль становить більшу частину цитоплазми (55 % від загального об'єму клітин), що не містить органел. Це колоїд, який складається зі складної суміші розчинених у воді органічних макромолекул - білків, жирів, вуглеводів та неорганічних речовин. Містить до 10000 різних видів білків, головним чином ферментів. У цитозолі знаходяться неорганічні (вода, солі, гази) і органічні речовини.
Неорганічні речовини. Вода є основною складовою частиною цитозолю. У середньому в клітинах міститься близько 75 % води. Завдяки своїм властивостям водне середовище забезпечує майже всі життєві процеси в клітинах. Зокрема, вода має такі властивості: 1) розчинник для речовин у клітині, внаслідок чого багато з них іонізуються водою, що полегшує хімічні реакції; 2) сприяє пересуванню речовин у клітині, із клітини в клітину в розчиненому стані; 3) ефективний термостабілізатор і зберігає тепло, утворене клітиною; 4) забезпечує постійний броунівський рух молекул.
Солі становлять 1-2 % цитозолю. У водному середовищі вони утворюють іони. Більшість солей клітин - це карбонати, бікарбонати, фосфати, сульфати і хлориди солей натрію, калію, кальцію, магнію та заліза. У першу чергу, вони відіграють істотну роль у підтримці осмотичності і кислотності цитозолю. Багато з них беруть участь у біологічних процесах і входять до складу деяких білків.
Хімічний склад і фізичний стан зумовлюють біологічні властивості цитозолю, що служать структурній цілісності і функціональній активності клітин. Насамперед - це підтримка метаболізму. Цитозоль - це середовище, де перебігають одночасно тисячі біохімічних реакцій. Вважається, що близько 70 % реакцій клітинного метаболізму відбувається в цитозолі, що містить тисячі різновидів ферментів. Це реакції гліколізу, глюконеогенезу, синтезу білків, жирних кислот, амінокислот, нуклеотидів та інші. На рибосомах у цитозолі синтезується багато білків, які використовуються клітиною для власних потреб. Рибосоми, зв'язані з ЕПС, утворюють білки на "експорт".
Функції органел клітини забезпечуються постійним, необхідним для них оточенням цитозолю. З цитозолю органели одержують необхідні речовини і викидають у нього відходи. Цитозоль бере участь у процесі підтримки гомеостазу клітини.
Реакції, що відбуваються у цитозолі, забезпечують сталість складу клітини та її структурної організації. У цитозолі постійно підтримується концентрація води, газів, субстратів хімічних реакцій, рН. Ці умови необхідні для перебігу біохімічних та фізіологічних процесів. Внаслідок постійного синтезу молекул (білків, амінокислот, нуклеотидів, вуглеводів, жирів та ін.) можливий обмін ушкоджених молекул на нові, синтезовані. Це стосується і постійної підтримки структури і складу всіх органел. Цитозоль є резервуаром різних субстратів (амінокислот, нуклеотидів, глюкози й інших), які постійно використовуються в обміні речовин для утворення нових структур або їх відновлення.
Цитоскелет. це сітка білкових фібрил і мікротрубочок, що вкривають зсередини цитоплазматичну мембрану і пронизують внутрішній простір клітини (рис. 3). Він характерний для всіх еукаріотичних клітин, а також є основним компонентом ворсинок і джгутиків найпростіших, хвостика сперматозоїдів, веретена поділу клітин. Цитоскелет складається з трьох типів структур: 1) мікротрубочки (найтовстіші), утворені кількома білковимифібрилами, які містять глобулярний білок - тубулін; 2) мікрофіламенти (найтонші), що мають здатність скорочуватися, утворюються глобулярним білком - актином; 3) проміжні філаменти (комбінація кількох мікрофіламентів).
Фібрили цитоскелета можуть за необхідності згруповуватися з мономерів білків і розпадатися після виконання функції. Мають здатність до скорочення і руху У клітині фібрили взаємодіють між собою за участі допоміжних білків. Вони вкривають з внутрішнього боку цитоплазматичну мембрану і пронизують внутрішній простір клітини. Цим досягається стабільність форми й об'єму клітини, а також можливість зміни форми, руху органел і клітини.

Рис. 3. Цитоскелет: 1 - клітинна мембрана; 2 - мітохондрія; 3 - комплекс Гольджі; 4 -рибосоми;5 - мікротрубочки; 6 - мікрофіламенти.
Функції цитоскелета.
1. Підтримка об'єму і форми клітин. Основну роль у цьому відіграє фібрилярна сітка, що вкриває зсередини мембрану (кортекс). Ця сітка спеціальним білком (онкерін) прикріплена до цитолеми. До цієї сітки приєднані нитки мікрофіламентів і мікротрубочок, що значною мірою стабілізує форму клітини.
2. Зміна форми клітин. Система білкових фібрил здатна до скорочення або розтягування. За рахунок цього може відбуватися зміна форми клітин (наприклад, формування псевдоподій у лейкоцитах).
3. Пересування органел і транспортних везикул. Фібрили цитоскелета прикріплені до клітинних органел. Це стабілізує їхнє положення в цитоплазмі. З іншого боку, зміна довжини фібрил призводить до переміщення клітинних структур.
4. Утворення мультиферментних компонентів. У місцях переплетення кількох фібрил цитоскелета створюються сприятливі умови для розміщення комплексу ферментативних білків. Це забезпечує структурну єдність ферментів та певний метаболічний процес.
5. Завдяки наявності щільної сітки мікрофібрилцитозоль набуває певної структури, що сприяє координованому розміщенню комплексів ферментів.
Цим досягається інтеграція всієї цитоплазми - об'єднання в єдине ціле.
6. Утворення веретена поділу під час мітозу. Веретено поділу утворене сіткою мікротрубочок, що "збираються" за участі центріоль і чітко впорядковано розташовуються в цитозолі.
7. Утворення ворсинок і джгутиків у найпростіших.
8. Утворення міжклітинних контактів (десмосом). Десмосоми - структури цитоплазматичних мембран, що належать одночасно двом сусіднім клітинам. Зв'язування клітин відбувається завдяки мікрофіламентам, що проникають через десмосому з однієї клітини в іншу.
9. Забезпечення скорочувальної функції м'язових волокон. Актинові філаменти є однією з головних частин скорочувального актиноміозинового комплексу.
Цитоплазма перебуває в постійному русі, чим забезпечується транспорт речовин до різних ділянок клітини. Рух цитоплазми – універсальній параметр всіх живих клітин. Він забезпечує структурну організацію живої клітини, її енергетичний обмін, веде до збалансованого розповсюдження попередників біосинтезу, продуктів біогенезу, впливає на мембранні потоки. Рух цитоплазми відбувається шляхом коливання, струменевого спрямування, колового руху, фонтануючого руху та ін.
Клітинні органели - диференційовані ділянки цитоплазми, що мають специфічний молекулярний склад. Це складні, високовпорядковані біологічні системи макромолекул, що утворюють певну просторову структуру, здатні до виконання спеціальних клітинних функцій. Клітини тварин містять багато внутрішньоклітинних мембран. Тому майже половина всього об'єму клітин укладена в окремі внутрішньоклітинні відсіки (компартменти), що називаються "органелами". Інший внутрішньоклітинний простір зайнятий цитозолем.
Класифікація органел. Клітинні органели умовно поділяють на мембранні, що оточені типовою біомембраною, і немембранні, що не мають такої оболонки.
Мембранні: 1) ендоплазматична сітка: а) зерниста; б) гладенька; 2) комплекс Гольджі; 3) лізосоми; 4) пероксисоми; 5) вакуолі; 6) мітохондрії; 7) пластиди (тільки в рослинних клітинах).
Немембранні: 1) рибосоми; 2) центріолі; 3) мікротрубочки; 4) мікрофіламенти.
Ендоплазматична сітка (ЕПС). ЕПС виявлена у всіх еукаріотичних клітинах, відсутня тільки в прокаріотів, у сперматозоїдах і зрілих еритроцитах. ЕПС утворена сіткою мембранних трубочок, цистерн і овальних везикул. ЕПС структурно зв'язана з оболонкою ядра. Розрізняють два типи ЕПС: гладеньку і зернисту, хоча вони структурно пов'язані між собою.
Зерниста ЕПС на своїй поверхні містить рибосоми, котрих немає на поверхні гладенької ЕПС. ЕПС утворює сітку мембранних каналів, що пронизують цитоплазму. Ендоплазматична сітка має значення в процесах внутрішньоклітинного обміну, оскільки збільшує площу внутрішніх поверхонь клітини, поділяє її на відсіки, що відрізняються за фізичним станом і хімічним складом, забезпечує ізоляцію ферментних систем, що, у свою чергу, необхідне для послідовного вступу в узгоджені реакції. Безпосереднім продовженням ендоплазматичної сітки є ядерна мембрана, що відмежовує ядро від цитоплазми, так і зовнішня мембрана (плазмолема), розташована на периферії клітини. Мембранні системи дуже лабільні і можуть змінюватися у залежності від фізіологічного стану клітини, характеру обміну, при рості та диференціюванні.
Цитоплазма еукаріотичних клітин містить мембранні шари, пухирці, трубочки, що відокремлюють у сукупності значний внутрішньоклітинний простір.
Функції ЕПС. Взаємозалежна система гладенької та зернистої ЕПС працює узгоджено і виконує ряд загальних інтегральних функцій: 1) мембрани ЕПС відокремлюють свій специфічний вміст від цитозолю; 2) у матриксі ЕПС відбувається нагромадження, збереження і модифікація синтезованих речовин; 3) ЕПС є важливою складовою системи внутрішньоклітинних мембран, забезпечує транспорт синтезованих речовин по внутрішніх порожнинах або за допомогою везикул у різні ділянки клітин; 4) структура ЕПС утворює велику мембранну поверхню всередині клітини, що важливо для багатьох метаболічних реакцій; 5) мембранна система пронизує всю клітину і виступає в якості "внутрішнього скелету".
Комплекс Гольджі (КГ), утворений комплексом із десятків сплощених дископодібних мембранних цистерн, мішечків, трубочок і везикул, у значній кількості зустрічається в секреторних клітинах. Внутрішній міжмембранний простір заповнений матриксом, що містить спеціальні ферменти. Електронно-мікроскопічні дослідження дозволили переконатися, що КГ збудований із мембран і нагадує стовпчик з порожніх дисків, накладених один на одного (рис 4).

Рис. 4. Комплекс Гольджі: 1 - зона формування; 2 - зона дозрівання
До його складу входить система трубочок із пухирцями на кінцях. Комплекс Гольджі має дві зони: зону формування, куди надходить синтезований матеріал із ЕПС за допомогою транспортних везикул, і зону дозрівання, де формується секрет і зрілі секреторні мішечки. До зони формування надходять синтезовані в ЕПС речовини, що знаходяться в мембранних везикулах. Вони зливаються з мембраною КГ, і вміст везикули надходить всередину комплексу. Речовини обробляються ферментами, після цього знову упаковуються у везикули і переносяться в зону дозрівання. У зоні дозрівання накопичується "дозрілий секрет", що відокремлюється у вигляді секреторних пухирців. У цьому компартменті утворюються також лізосоми і пероксисоми.
Функції комплексу Гольджі: 1) нагромадження і модифікація синтезованих макромолекул; 2) утворення складних секретів і секреторних везикул; 3) синтез і модифікація вуглеводів, утворення глікопротеїдів; 4) КГ відіграє важливу роль у відновленні цитоплазматичної мембрани шляхом утворення мембранних везикул і наступного злиття з клітинною оболонкою; 5) утворення лізосом; 6) утворення пероксисом.
Лізосоми - це невеликі (0,2-0,8 мкм), вкриті мембраною, круглі тільця (рис. 5). Зустрічаються вони у всіх клітинах рослин і тварин, можуть локалізуватися в будь-якому місці клітини.
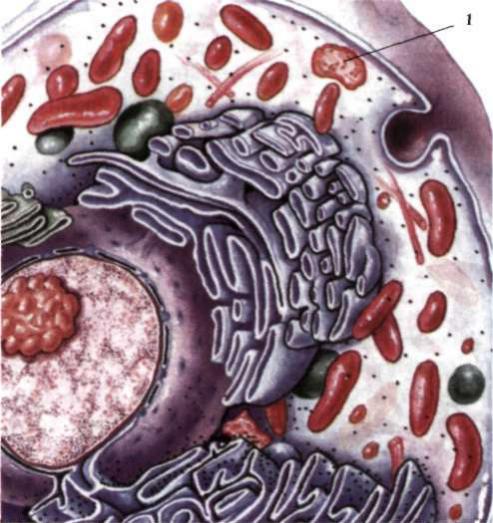
Рис. 5. Клітина еукаріотів: 1 - лізосома.
Вміст лізосом складають різні класи гідролітичних ферментів, наприклад, протеази, нуклеази, ліпази, фосфоліпази та ін. Всього нараховується до 40 різних ферментів. Ці ферменти руйнують великі молекули складних органічних сполук, що надходять до клітини (білки, нуклеїнові кислоти, полісахариди). У лізосомах зазнають руйнації мікроорганізми і віруси. Ферменти лізосом перетравлюють зруйновані структури або цілі клітини. Ці процеси називаються аутофагією (від грец. αυτός - самий, φάγος - пожирання).
Кожна лізосома вкрита щільною мембраною, що ізолює ферменти від цитоплазми. Ушкодження мембран лізосом і вихід із них у цитоплазму ферментів викликає швидке розчинення (лізис) клітини.
Лізосоми різноманітні за своєю природою і можуть утворюватися різними шляхами. У кожному випадку формуються морфологічно різноманітні лізосоми, що розщеплюють матеріал із різних джерел. У центрі цих шляхів знаходиться "проміжний компартмент" - ендолізосома. Процес перетравлення лізосомними ферментами об'єктів, що надходять до клітини шляхом фагоцитозу, відбувається у вакуолях, які називаються фагосомами. Продукти перетравлення потрапляють у цитоплазму, а неперетравлений матеріал залишається у фагосомах і зменшується в розмірах. Такі структури називаються залишковими тільцями.
Пероксисоми - маленькі сферичні тільця, вкриті мембраною (рис. 6).
Виявляються майже у всіх клітинах еукаріотів. їх діаметр становить 0,3-1,0 мкм, утворюються в комплексі Гольджі. Пероксисоми містять в основному ферменти для руйнації пероксиду водню. Пероксид водню, що утворюється в результаті окиснення деяких органічних речовин, є токсичним для клітини і тому негайно руйнується каталазою пероксисоми:
2H202 -> 2Н20 + 02
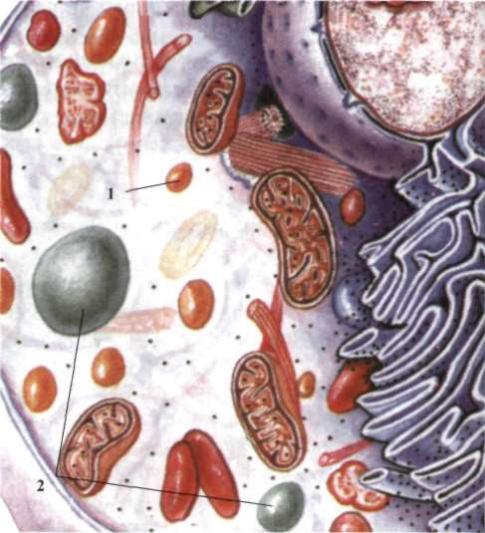
Рис. 6. Клітина еукаріотів: 1 - пероксисома, 2 - вакуолі.
Пероксисоми беруть участь у процесі β-окиснення жирних кислот. До 50 % жирних кислот руйнуються в пероксисомах. Вони містять також й інші окисні ферменти.
Вакуолі - це порожнини в цитоплазмі, оточені мембраною та заповнені рідиною. В еукаріотичних клітинах є різні типи вакуоль. Вакуолі можуть виникати з пухирців, які відокремлюються від ендоплазматичної сітки, або комплексу Гольджі. Вони заповнені водним розчином органічних і неорганічних сполук, серед них – продуктів обміну або пігментів. Функції вакуоль різноманітні: вони підтримують тургорний тиск, зберігають поживні речовини і накопичують продукти обміну. Скоротливі вакуолі одноклітинних тварин регулюють осмотичний тиск у клітині, беруть участь у виведенні продуктів обміну, а також сприяють надходженню в клітину води.
Мітохондрії (від грец. μίτος - нитка, χονδράς - зернятко) - це органели, в яких енергія хімічних зв'язків органічних речовин перетворюється на енергію фосфатних зв'язків АТФ. Мітохондрії досить великі овальні органели (0,2-2,0 мкм), вкриті двома мембранами. Вони зустрічаються майже в усіх еукаріотичних клітинах, за винятком анаеробних найпростіших і еритроцитів. Мітохондрії хаотично розподілені по цитоплазмі, хоча частіше виявляються біля ядра або в місцях із високими потребами енергії. У м'язових клітинах вони розташовані між міофібрилами. Органели можуть змінювати свою структуру і форму, здатні переміщуватися всередині клітини. Кількість мітохондрій може змінюватися залежно від активності клітини від кількох десятків до кількох тисяч.
Органела містить зовнішню і внутрішню мембрани з вузьким міжмембранним простором (рис. 7).
Внутрішня мембрана утворює численні вирости - кристи, що оточені матриксом, в якому знаходиться багато ферментів, рибосоми, одна молекула ДНК.
Мітохондрії - органели розміром з бактерію, що використовують енергію окиснення для утворення АТФ.
Зовнішня мембрана легко проникна для багатьох невеликих молекул. Містить ферменти, що перетворюють речовини на реакційноздатні субстрати, бере участь в утворенні міжмембранного простору.

Рис. 7. Клітина еукаріотів: 1 - мітохондрія; 2 - зовнішня мембрана; 3 – внутрішня мембрана; 4 - криста; 5 - матрикс; 6 - рибосома; 7 – між мембранний простір.
Внутрішня мембрана погано проникна для більшості речовин. Вона утворює вирости – кристи всередині матриксу. Ця мембрана містить ферменти, що беруть участь у наступних важливих процесах:
а) ферменти, що каталізують окисно-відновні реакції дихального ланцюга і транспорту електронів. В результаті утворюється надлишок Н+ у міжмембранному просторі;
б) специфічні транспортні білки беруть участь в утворенні градієнту Н+;
в) ферментний комплекс АТФ-синтетази, що синтезує АТФ.
Міжмембранний простір використовується для градієнта іонів Н+ на внутрішній мембрані, що є необхідною умовою синтезу АТФ.
Матрикс - це простір мітохондрії, обмежений внутрішньою мембраною. Він утворений сотнями різних ферментів, що беруть участь у руйнації органічних речовин до С02 і Н20. При цьому вивільняється енергія хімічних зв'язків між атомами молекул органічних речовин і перетворюється в макроергічні зв'язки АТФ. У матриксі знаходяться рибосоми і молекула мітохондріальної ДНК. Рибосоми мітохондрій і ДНК забезпечують синтез необхідних органелі білків. Основна інтегральна функція мітохондрій - перетворення енергії та утворення АТФ, що містить макроергічні зв'язки.
Особливості мітохондрій, що вказують на їхню подібність із прокаріотами, розглядають як доказ симбіотичного походження цієї органели. Згідно з такою гіпотезою, деякі аеробні прокаріоти проникли в більшу анаеробну клітину. Можливо, спочатку вони вели паразитичний спосіб життя. Надалі партнери цього співжиття в процесі еволюції пристосувалися один до одного і колишній "паразит" перетворився в органелу, необхідну для існування клітини. Але як органели предки мітохондрії загубили частину свого генетичного матеріалу. В еукаріотичних клітинах мітохондріальна ДНК кодує лише частину мітохондріальних білків, більша ж кількість їх синтезується поза мітохондріями і пов'язана з ядерною ДНК.
Пластиди - двомембранні органели клітин рослин і деяких тварин (джгутикових). У клітинах вищих рослин розрізняють три типи пластид: хлоропласти, хромопласти та лейкопласти.
Хлоропласти - забарвлені у зелений колір завдяки пігменту хлорофілу. Між зовнішньою та внутрішньою мембранами хлоропластів є міжмембранний простір завширшки близько 20-30 нм. Внутрішня мембрана утворює вгини - ламели та тилакоїди.
Ламели мають вигляд плоских видовжених складок, а тилакоїди - сплощених вакуоль або мішечків. Ламели утворюють сітку розгалужених канальців. Між ламелами розміщені тилакоїди, зібрані у вигляді стопки монет (грани). У тилакоїдах знаходяться фотосинтетичні пігменти - хлорофіл, каротиноїди та ферменти, які потрібні для здійснення різноманітних біохімічних процесів У матриксі пластид є також власний білоксинтезувальний апарат (молекули ДНК і рибосоми). Основна функція хлоропластів - фотосинтез.
Лейкопласти - безбарвні пластиди, які відрізняються від хлоропластів відсутністю розвиненої ламелярної системи. Вони забезпечують синтез і гідроліз крохмалю і білків.
Хромопласти - пластиди, які надають забарвлення (жовтого, червоного та ін.) пелюсткам, плодам, листкам. Забарвлення хромопластів зумовлюють пігменти - каротиноїди. Внутрішня мембранна система у хромопластів відсутня або утворена поодинокими тилакоїдами.
Рибосоми (від лат. ribes - потік, струмінь і грец. σώμα - тіло) - невеликі грануло подібні сферичні тільця, розміром від 15 до 35 нм (рис. 8). Рибосоми складаються із двох субодиниць, розташовані в цитоплазматичному матриксі або зв'язані з мембранами ендоплазматичної сітки.
Субодиниці рибосом утворюються в ядерці, а потім через ядерні пори окремо одна від одної надходять до цитоплазми. їх кількість у цитоплазмі залежить від синтетичної активності клітини і може складати від сотні до кількох тисяч на одну клітину, їх функцією є синтез білків. Найбільша кількість рибосом виявлена в клітинах, що інтенсивно синтезують білки. Ці органели зустрічаються також у мітохондріальному матриксі й хлоропластах. Рибосоми будь-яких організмів - від бактерій до ссавців - характеризуються подібністю структури і складу. Кожна субодиниця складається з кількох різновидів молекул рРНК і десятків різновидів білків, приблизно в однаковій пропорції. Маленька і велика субодиниці знаходяться в цитоплазмі окремо одна від одної, доки не беруть участі в білковому синтезі.
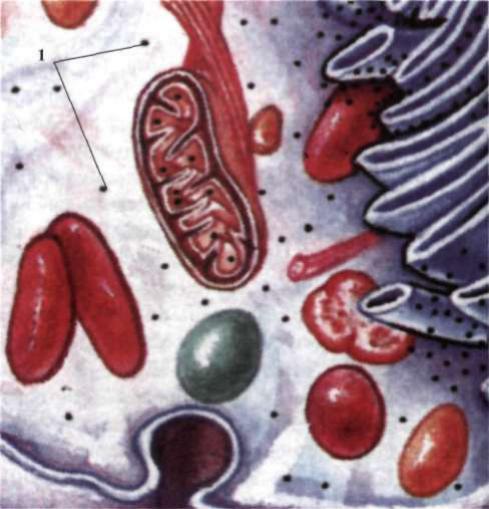
Рис. 8. Клітина еукаріотів: 1 – рибосоми
Вони об'єднуються одна з одною і з молекулою іРНК за необхідності синтезу і знову роз'єднуються з припиненням процесу. Якщо з однією молекулою іРНК з'єднуються кілька рибосом, то утворюються полісоми, що містять від 5 до 70 рибосом.
Клітинний центр (центросома) - органела, що складається з двох дрібних утворень: центріоль і променевої сфери навколо них (рис. 9).
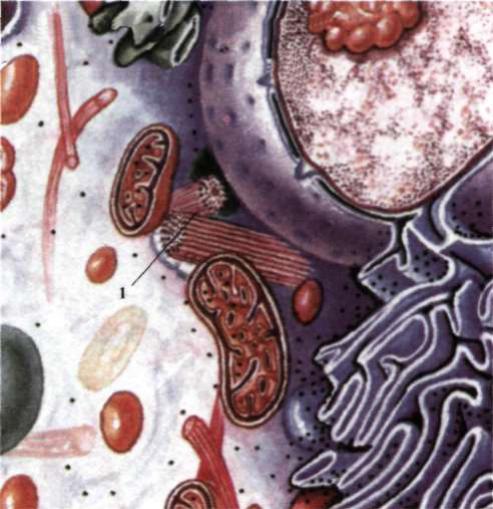
Рис. 9. Клітина еукаріотів: 1 - центроєома.
За допомогоюелектронного мікроскопа встановлено, що кожна центріоля - це циліндричне тільце довжиною 0,3-0,5 мкм і діаметром близько 0,15 мкм. Стінки циліндра складаються з 9 пар паралельно розташованих мікротрубочок, що утворені білками. Центріолі розміщуються перпендикулярно одна до одної. Клітинний центр іноді займає геометричний центр клітини (звідси назва органели). Частіше ж він відтиснутий ядром або включеннями до периферії, але обов'язково розташовується поблизу ядра на одній осі з центром ядра і клітини. Активна роль клітинного центра виявляється при поділі клітини.
Центріолі подвоюються і розходячись у протилежні боки, формують полюси клітини, що ділиться. Зазначені структури утворюють веретено поділу.
Центріолі беруть участь в утворенні мікротрубочок цитоскелета. Вони також формують базальне тіло, що лежить в основі джгутиків.
Мікротрубочки і мікрофіламенти - це немембранні органели, які побудовані з скоротливих білків (тубуліну, актину, міозину тощо). Мікротрубочки (рис. 10) циліндричної форми, порожнисті, діаметром 10-25 нм. Вони беруть участь у формуванні веретена поділу, у внутрішньоклітинному транспорті речовин, входять до складу війок, джгутиків, центріоль.

Рис. 10. Клітина еукаріотів: 1 - мікротрубочки
Мікрофіламенти причетні до формування цитоскелета клітини. Розташовані ці органели під плазматичною мембраною. Пучечки мікрофіламентів одним кінцем прикріпляються до мембрани, а іншим - до різних органел, молекул біополімерів. Вони беруть участь у зміні форми клітини, наприклад, під час її руху. У м'язових клітинах пучечки мікрофіламентів розміщені вздовж їхньої осі (волокна актину та міозину).
Ядро було відкрито Броуном в 1831 р. Воно є обов'язковим і найважливішим компонентом клітки. Йому належить головна роль в збереженні, передачі і реалізації спадкової інформації. Ядро клітки, що не ділиться, називається інтерфазним. В цей час воно має сферичну або овальну форму і в недиференційованій клітині розташовується в центральній частині. Ядро оточено ядерною оболонкою (мембраною), містить каріоплазму, ядерце і хроматин (рис.11,12). зосереджений в ядрі (від грец. xapuov - ядро, лат. nucleus - ядро). Ядерна оболонка відокремлює генетичний матеріал і молекулярно-генетичні процеси від цитоплазми, забезпечує автономність і незалежність спадкових механізмів. Більшість клітин містить тільки одне ядро. При виділенні ядра клітина не може довго існувати, так само як і ядро, виділене з клітини, гине. Ядро звичайно локалізується в центрі клітини.
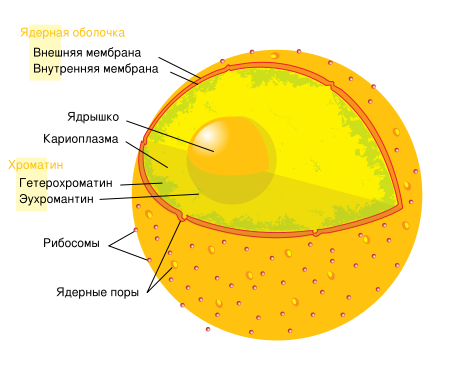
Рис. 11. Будова ядра клітини.
.

Рис. 12. Будова ядра: 1 - ядерна оболонка; 2 - зовнішня мембрана; 3 – внутрішня мембрана; 4 - перинуклеарний простір; 5 - каріоплазма; 6 - хроматин; 7 - ядерце; 8 - ядерні пор
Структура ядра. Ядро складається з декількох компонентів, що виконують різні функції: ядерна оболонка, каріоплазма, хроматин, ядерце.
Ядро круглої, кулеподібної форми. У клітинах з високою фізіологічною активністю форма ядер складна, що збільшує співвідношення поверхні ядра до його об'єму. Співвідношення об'ємів ядра і цитоплазми називається ядерноцитоплазматичним співвідношенням. Зміна цього є одним із чинників клітинного поділу або порушення обміну речовин.
Хімічний склад. До складу сухої речовини ядер входить - 80 % білків, 12 % ДНК, 5% РНК, 3% ліпідів і деяка кількість Mg2+. Більшість білків - ферменти, що каталізують молекулярногенетичні процеси. Крім цього, гістонові й негістонові білки разом із ДНК утворюють хроматин. Певна частина білків складає основу ядра у вигляді мікрофіламентів і ядерної пластинки - сітки білкових ниток, що вистилає внутрішню поверхню ядра.
Спеціальні білки зв'язуються з РНК і утворюють субодиниці рибосом. Деякі білки входять до складу ядерних пор.
В ядрі є три різновиди РНК: іРНК, тРНК, рРНК.
Каріоплазма. Каріоплазма містить велику кількість води (75-80 %), в якій сконцентровані: хроматин (гетерохроматин і еухроматин), мікрофіламенти, ядерце, ферменти.
Ядерна оболонка. Ядерна оболонка вкриває ядро, і має специфічний хімічний склад, який сприяє перебігу важливих молекулярно-генетичних процесів. Складається із зовнішньої і внутрішньої мембран; між ними знаходиться перинуклеарний простір, який через канали ендоплазматичної, сітки зв'язаний з різними ділянками цитоплазми. Обидві мембрани пронизані численними порами. Через них відбувається вибірковий обмін речовин між ядерним вмістом і цитоплазмою. Всередину ядра надходять білки, АТФ, нуклеотиди, а з ядра в цитоплазму виходять субодиниці рибосом, тРНК та іРНК. При розподілі клітини ядерна оболонка кожного разу розкладається на окремі мембранні пухирці - везикули - і знову збирається в дочірніх клітинах, оточуючи генетичний матеріал. Вважається, що ядерна оболонка еукаріотів утворилася з мембран ЕПС.
Обидві мембрани типової будови – ліпідний бішар з вбудованими в нього білками. Зсередини ядерна оболонка вкрита білковою сіткою – ядерною ламіною, що зумовлює форму й об'єм ядра.
До ядерної ламіни теломерними ділянками приєднані нитки хроматину. Мікрофіламенти утворюють внутрішню "основу" ядра. Вони підтримують його форму, а також слугують місцем прикріплення хроматину. Внутрішній "скелет" ядра має велике значення для забезпечення упорядкованого перебігу основних процесів транскрипції, реплікації, процесингу.
Зовні ядро також вкрите мембраною, яка є елементам цитоскелета клітини. Зовнішня мембрана може мати на своїй поверхні рибосоми і з'єднана з мембранами ендоплазматичної сітки.
Ядерна оболонка має властивість вибіркової проникності. Потоки речовини регулюються специфічними властивостями білків мембран і ядерних йор.
Основні функції оболонки: 1) створення компартмента клітини, де сконцентрований генетичний матеріал і умови для його збереження і подвоєння; 2) відокремлення від цитоплазми, в якій інакший вміст речовин і проходять інші процеси; 3) підтримання форми й об'єму ядра, або їх змін; 4) регуляція потоків речовин всередину і назовні ядра. З ядра крізь пори в цитоплазму надходять різні види РНК і субодиниці рибосом, а всередину ядра переносяться необхідні білки, вода, іони тощо.
Функції ядра: 1) збереження спадкової інформації в молекулах ДНК; 2) реалізація спадкової інформації шляхом регуляції синтезу білків. Завдяки цьому підтримується структурна впорядкованість клітин, регулюються їх метаболізм, функції та процеси поділу; 3) передача спадкової інформації наступним поколінням внаслідок реплікації ДНК шляхом утворення хромосом та їх поділу.
Найважливіші молекулярно-генетичні процеси, що відбуваються в ядрі: реплікація ДНК, транскрипція всіх видів РНК, процесинг, утворення рибосом.