

25.Симбиотические отношения.
Симбиоз-отношения м\у орг-ми, обычно относящ-ся к разным видам и нах-мися в более или менее длит. контакте, при кот. оба орг-зма извлекают из этих отношений пользу и ни один не испытывает ущерба.
Комменсализм – это такая форма взаимоотношений м/у 2 видами, когда деят-ть 1 из них доставляет пищу или убежище другому или одностороннее ∆ одного вида другими без принесения ему вреда. Комменсализм основанный на потребление остатков пищи хозяев, называется нахлебничеством. (Львы и гиены, они подъедают недоеденные львами остатки). Квартиранство –∆ убежища либо в постройках, либо в телах других видов (молодь рыб прячется под стрекательными нитями медуз). Нидиколы- постоянные норовые или гнездовые сожители.
Н-р: растения, прикрепленные к поверхности других растений, кот.поддерживают их в кач-ве споры, наз-ся эпифитами. К ним отн-ся растения, живущие на др.раст-ях или в их отмерших внеш тканях, но не поглощающие воду и эл-ты мин.питания из живых органов. В троп.лесах эпифиты – цветковые раст-я(орхидные). Хозяин, на котором селятся называетя форафитом.
Среди животных комменсалы: рыбы-прилипалы, рыбы-лоцманы, насекомые, поселяющ-ся в норах млекоп-х, гнездах птиц.
Мутуализм – взаимовыгодное отнош-е видов. Мутуалистич-е связи м. возникать на основе предшествующего паразитизма или комменсализма. Ст. развития взаимовыгодного сож-ва м.б.различной – от временных, необязательных контактов до состояния, когда присутсвие партнёра становится обязательным усл-ем жизни каждого из них. Такие неразделимые полезные связи 2 видов наз-ся симбиоз. Н-р: лиш-ки – тесное сожительство гриба и водорослей, и сож-ва многих видов дер-ев с микоризными грибами, бобовые растения – с клубеньковыми бактериями, симбиоз ВР с гриб, бактериями., водорослями, симбиоз ВР с грибами, образующими экто- и эндомикоризу. Для всех типов микориз доказано, что грибной симбионт улучшает обеспечение растения фосфором.
Виды микосимбиотических отношений:
1) чехольчатая энтомикориза – грибы передают растению влагу, минеральные вещества (особенно фосфор), а растение поставлет грибу углеводы. Гриб выделяет вещества, препятствующие заражению.
2) везикулярно-арбускулярная эндомикориза – гифы гриба входят в ткань растений. Растения способны перерабатывать гифы гриба. Даже можно отнести к паразитизму, т.е. двойной паразитизм.
3) вересково-арбутоидная энтомикориза – характерна для вида семейства вересковых. У вересковых характерно усвоение органических веществ благодаря грибному партнеру. Вересковые часто селятся на болотах, где много органики. Грибы помогают перерабатывать её.
4) миксимикотрофия орхидных – на стадии образования заростка орхидные паразитируют на грибах, т.к. их семена очень маленькие и мелкие, питат. веществ мало.
Больш.роль в функ-ии экосис-м и биосф играют симб.отнош-я м\у ВР и азотфиксир бактериями, вовлекающими молекулярн. азот в биологич.круговорот – бактериосимбиотрофия. У простейших, например, роль передвигательных органов могут играть другие простейшие, т.е. как реснички быть и помогать передвигаться.
К взаимовыгодн. отнош-ям относ. отношения ВР с насекомыми-опылителями.
Симбиот.отнош-я у жив-х:благодаря наличию в пищ.тракте жгутиковых термиты способны use в пищу древесину, выполняя важнейш.ф-цию в ЭС по минерализации целлюлозы. Взаимоотношения м\у раком-отшельником и актинией-тоже взаимовыгодние.
+симбиоз однокл.водорослей водорослей с грибами, простейшими, моллюсками.
26 ХИЩНИЧЕСТВО. ПОПУЛЯЦИОННЫЕ СТРАТЕГИИ Ф-РЫ СТАБИЛЬНОСТИ Х И Ж.
Хищ-во –взаимодействие 2х поп, «-» сказывающих на росте и выживании одной поп, умерщвляя ее. Особенность «-»вз-дей-я поп-й закл-ся в том, что при их синхронной эволюции в стабильной ЭС степень «-» влияния ↓, Естес.отбор(ЕО) стремится снизить «-» влияние, поскольку сильное влияние м. унич-ть 1 поп или обе. Жесткая конк-я чаще отмеч-ся там, где контакт м/у поп-ми установился недавно. Х- м.б. полифаги (универсалы, поедают многие виды), олигофаги (некот виды), монофаги - специалисты (только 1 вид-ех:коршун-слизниед), м.б. эф-ми и нет. Х-специалист тратит много времени на обработку и мало на поиск (львы). Универсалы наоборот.
Опыт Гаузе: изуч взаим-е м\д 2идами инфузорий: туфелька(Paramecium caudatum)-Ж, дидиниум(Didinium)-Х: сначала нач-ет ↑ чис-ть Ж, затем Х, затем активно выедает и Ж не остается, затем ↓ чис-ть Х до 0 – сист не устойчива. Гаузе в пробирку внес стекл.вату, внутри α м. прятаться Ж(Х туда попасть не м.)- Х будет выедать всех, кто не спрятался, затем погибнет сам, а спрятавшиеся попул-я Ж будет ↑-ся, за счет спрят-ся особей.Наиб-го равновесия Гаузе добил-ся переодич-ки внося в пробирку Ж и Х.
Реакция хищника на плотность популяции жертвы:
1)ф-циональная (учит.скор.потреб.пищи отдельной особи и Х в завис.от плотности попул.Ж). 2)Числ-я. Хар-ризуется ∆ числа Х путем миграции и размножения (аномальное увелич. численности Х при увеличении численности Ж).
По оси x - плотность популяции жертвы, по оси у – доля популяции жертвы.
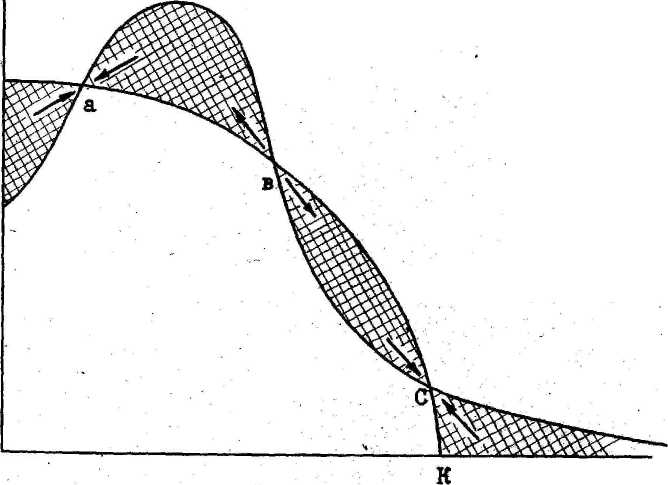 Ф-ры,
обеспечивающие стабильность сис-мы
“Х-Ж”. ф-ры,
обеспеч-щих стабильное устойчивое
равновесие в отн-о-ях Х и его Ж: 1.
Неэффективность хищника (или бегство
жертвы). К неэффективным хищникам относят
волк, лев, тигр. Как видно из модели,
сис-ма “Х-Ж”
достигает
одной из двух возможных точек равновесия.
Положение первой (с) определяется
емкостью данной среды для данной Ж в
отсутствие Х, положение второй (a)
определяется способностью Ж при низкой
численности популяции находить убежища.
Точка равновесия в сис-ме устанавливается
в зависимости от соотношения эффективности
Х как охотника и потенциала роста
популяции Ж.
Неэфф-е
Х уничтожают наиболее уязвимую часть
популяции Ж, доводя ее до уровня емкости
среды (точка (с)). При этом регуляция
численности Ж целиком зависит от реакции
ее на пищевые ресурсы. Эффективные Х
доводят популяцию Ж до точки равновесия
(a).
Однако вблизи этой точки даже для Х
этого типа эфф-сть выедания падает, так
как число сохранившихся особей жертвы
становится меньше, они удалены друг от
друга, часть их имеет доступ к надежным
убежищам.
Ф-ры,
обеспечивающие стабильность сис-мы
“Х-Ж”. ф-ры,
обеспеч-щих стабильное устойчивое
равновесие в отн-о-ях Х и его Ж: 1.
Неэффективность хищника (или бегство
жертвы). К неэффективным хищникам относят
волк, лев, тигр. Как видно из модели,
сис-ма “Х-Ж”
достигает
одной из двух возможных точек равновесия.
Положение первой (с) определяется
емкостью данной среды для данной Ж в
отсутствие Х, положение второй (a)
определяется способностью Ж при низкой
численности популяции находить убежища.
Точка равновесия в сис-ме устанавливается
в зависимости от соотношения эффективности
Х как охотника и потенциала роста
популяции Ж.
Неэфф-е
Х уничтожают наиболее уязвимую часть
популяции Ж, доводя ее до уровня емкости
среды (точка (с)). При этом регуляция
численности Ж целиком зависит от реакции
ее на пищевые ресурсы. Эффективные Х
доводят популяцию Ж до точки равновесия
(a).
Однако вблизи этой точки даже для Х
этого типа эфф-сть выедания падает, так
как число сохранившихся особей жертвы
становится меньше, они удалены друг от
друга, часть их имеет доступ к надежным
убежищам.
Чернов:Порог ценность корм пятна(КП): opt времени пребыв-я в КП устанавл-ся V-ю получ-я Е при выходе из П. Х, доб-щий пищу, max-рует V получения Е. Е пост-ет неравн-но, т.к. пища представл.в виде пятен и в период перехода м\д пятнами Е не поступает↓(Vскор=совокЕ/t). Х б нах-ся до тех пор, пока не начнет уменьшатся скорость получ-я Е, чем меньше расстояние до след кормового пятна, тем раньше он его покинет.
2. Эко ограничения, налагаемые внеш средой на популяцию. Точка равновесия зависит от внеш ф-ов, огранич-х рост Ж. Ф-ры среды ограничивают и рост численности Х. Верх точка равновесия во взаимоотношениях Х-Ж зависит от ограничений, налагаемых на популяцию Ж внеш средой. Регуляция числ-ти попу-ции Х внеш ф-ми, обусловл-я огранич числом мест для размножения, недостатком воды и Альтер-х пищ ресурсов, снижает давление, оказываемое на популяцию Ж Х, и повышает регулирующее возд-е внеш ф-ров.
3. Наличие у Х альтерн пищ ресурсов. В случае недостаточного кол-ва или низкой активности Ж Х временно использует другие источники пищи- позволяет ему эффективно регулировать численность Ж.
4.Уменьшение запаздывания в реакции Х на рост популяции Ж приводят к затуханию колебаний численности популяций и повышает общую устойчивость сис-мы Х – Ж вблизи любой из точек равновесия.
Уравнение Лотки-Волтерры:
![]() ,
,
![]()
r1 – скорость увеличения популяции жертвы, λ1 – вероятность того, что при встрече с хищником жертва будет съедена, λ2 – коэффициент хищничества («доход», полученный хищником при поедании жертвы), β – смертность хищника.
Ф-ры, опред-е стабильность взаим-я: не эф-сть Х, не однородность среды в пространстве и во времени, ограничение ф-рами среды (иначе при однород среды Х истребит все), спос-ть Х переключатся на альтер-ые ресурсы.
Стратегии: если Х неэфект-ый – Ж долгоживущая, имеет способы ухода от Х, и Х исп-ет старых и больных, основ-я часть Ж не затрагив-ся. если эффективный Х, то Ж должна иметь выс скорость роста попул, иметь короткий жизн цикл, ост-ся только размножаться.
Паразити́зм. Коадаптация паразита и хозяина.
Паразити́зм— один из видов сосуществования организмов. Это явление, при котором два и более организма, не связанные между собой филогенетически, генетически разнородны, сосуществуют в течение продолжительного периода времени, при этом они находятся в антагонистических отношениях. Паразит использует хозяина как источник питания, среду обитания.
Формы паразитизма и связанные с этим взаимные адаптации паразитов и их хозяев чрезвычайно многообразны. Различают эктопаразитизм, при котором паразит обитает на хозяине и связан с его покровами (клещи, блохи, вши и др.), и эндопаразитизм, при котором паразит живет в теле хозяина (паразитические черви, простейшие и др.) По степени тесноты связей паразита и хозяина выделяют две формы паразитизма: облигатный и факультативный. В первом случае вид ведет только паразитический образ жизни и не выживает без связи с хозяином (паразитические черви, вши). Факультативные паразиты, как правило, ведут свободный образ жизни и лишь при особых условиях переходят к паразитическому состоянию.
Организмы-хозяева обычно в ходе эволюции также изменяются; у них появляются защитные механизмы против паразитов. Растения часто продуцируют токсины, которые могут одновременно наносить эффективное воздействие на паразитических грибов, бактерий и растительноядных животных. Иммунная система позвоночных способна нейтрализовывать большинство паразитов (особенно микропаразитов) при их контакте с жидкостями организма.
С другой стороны, большинство паразитов, особенно микроорганизмов, эволюционно приобрели адаптации для защиты от противодействия хозяев. Такими адаптациями могут быть утолщенные клеточные стенки бактерий, клещи-захваты на конечностях, предотвращающие вычесывание блох и клещей из волосяного покрова, и т. д.
При описанном типе взаимодействия оба вида (хозяин и паразит) обычно совместно эволюционируют к более-менее стабильному состоянию, когда их влияние на численность друг друга становится минимально возможным
Морфофизиологические адаптации связаны с изменением внешнего и внутреннего строения паразитов и функционирования их систем органов. Они подразделяются на:
- прогрессивные адаптации: наличие органов фиксации (присоски, крючья, коготки вшей, ротовой аппарат клещей); сложное строение наружных покровов (кутикула, тегумент); молекулярная "мимикрия" (сходство структуры белков и ферментов паразита и хозяина); выделение кишечными паразитами антиферментов (зашита от переваривания соками хозяина); внутриклеточное паразитирование; иммунносупрессивное действие паразитов (эндопаразиты секретируют протеазы, разрушающие иммунные комплексы и клетки хозяина) и др.
- регрессивные: редукция органов движения и некоторых систем (кровеносной, дыхательной); упрощение строения нервной системы и органов чувств.
Паразиты являются естественными компонентами биогеоценозов, и вызываемые ими болезни и эпизоотии принадлежат к числу нормальных функций биоценоза. "Вредный" для одной особи паразит становится биологически "полезным" для популяции и вида. Паразиты являются более совершенными регуляторами численности популяций животных, чем хищники, потому что они регулируют численность и самих хищников. Популяция хозяев, лишившаяся паразитов как эффективного механизма регуляции собственной численности, будет обречена на гибель. "Роль паразитов в природе - стабилизация экосистем" (В.Н. Беклемишев). Подобное значение паразитов ни в коей мере не распространяется на современные человеческие популяции.
Нарушение равновесия в отношениях паразит – хозяин вследствие деятельности человека.
Влияние человека на отношения «паразит - хозяин». В сельскохозяйственных экосистемах заражение скота паразитами, к которым у животных нет иммунитета, может привести к гибели многих животных. Представляют опасность взаимоотношения паразитов и человека, который может заболевать гельминтозами, вызываемыми разными видами глистов, лямблиозом (при заражении простейшим - лямблией), болезнями бактериальной и вирусной природы. Отсутствие иммунитета к грибам-паразитам характерно для многих видов культурных растений. Наиболее распространенными патогенами из числа этих паразитов являются виды, вызывающие ржавчину, которая поражает хлебные злаки и многие плодовые культуры - виноградную лозу, яблоню и др. В большинстве случаев человек вынужден компенсировать отсутствие иммунитета у растений использованием специальных препаратов-фунгицидов, которые достаточно опасны для окружающей среды. Современные методы генетики и селекции позволяют вести отбор на повышение иммунитета к патогенам. К примеру, устойчив к грибным заболеваниям гибрид пшеницы и ржи (тритикале). Особенно эффективны методы генной инженерии (создание генетически модифицированных растений). Катастрофическими бывают последствия заноса человеком паразитов в новые районы, где у их потенциальных хозяев отсутствуют механизмы защиты. В ХХ в. произошли ботанические катастрофы. В Америке погиб зубчатый каштан - Castanea dentata от занесенного из Китая паразитического гриба Endothia parasitica, вызывающего «рак каштана» (заражение произошло от китайского каштана, который был использован для создания гибрида с местным видом). В Европе от «голландской болезни», вызываемой грибом Ophiostoma ulmi, который переносится жуком- короедом, почти полностью исчез вяз. К еще более серьезным последствиям привело распространение вируса «коровьей чумы» в саваннах Африки. Из-за этого вируса, который первоначально вызвал болезни у домашнего скота, а затем поразил многие виды диких крупных копытных животных, резко снизилась интенсивность поедания фитофагами растительности саванны, и в первую очередь ее древеснокустарникового компонента, что сказалось на его отношениях с травяным покровом саванны. Кустарники и деревья стали бурно разрастаться, подавляя травы, что увеличило количество пожаров, которые при таком сомкнутом древесном пологе стали более частыми и верховыми, охватывающими не только травяной ярус, но и деревья. После пожаров деревья отрастали плохо и замещались кустарниками, корневища которых позволяли им сохраняться во время пожара. Только в 1970-е годы, когда удалось снять влияние на экосистемы вируса «коровьей чумы», вылечив от болезни домашний скот, поставлявший паразита популяциям диких животных через выделения слюны на пастбищах, где их выпас чередовался, процесс был остановлен. Восстановились плотность популяций животных, питающихся ветками, и, баланс между древеснокустарниковым и травяным компонентами экосистемы саванны, которая приобрела первозданный облик. Впрочем, в некоторых случаях популяции за несколько поколений сами вырабатывают иммунитет к новому для них паразиту. Так случилось с кроликом, который был интродуцирован из Европы в Австралию (см. 10.4). Аналогично протекают самые тяжелые эпидемии у человека, что иллюстрирует история массового заболевания чумой, которая распространялась блохами, обитающими на крысах. В начале эпидемии она буквально косила людей, и число заболевших и умерших было очень велико, но дальше смертность снижалась, так как незаболевшие уже имели иммунитет, а у переживших болезнь он формировался. По этой причине спустя всего несколько лет эпидемии быстро шли на спад.
Любые количественные изменения в составляющих устойчивости паразитарных систем ведут к колебаниям уровней, нарушающих устойчивость. В нетрансформированных биоценозах нарушения восстанавливаются за счет саморегуляции. Но в условиях антропопрессии они могут приводить к постоянной или временной утрате устойчивости, т.е.к разбалансированию паразитарных систем. Из этого следует важный вывод,что в условиях антропопрессии (например, в урбанизированных экосистемах), практически любая паразитарная система, вне зависимости оттого, участвует в ней человек как хозяин конкретного вида паразита или нет,должна рассматриваться и оцениваться в комплексе с социальными факторами и разнообразными загрязнениями, т.е. "вместе с человеком". Онне только как биологический хозяин паразитов (им он может быть, а может ине быть…), но и как "преобразователь природы", своей деятельностью,нарушает многие важнейшие функциональные связи, сложившиеся внутриконтуров огромного числа паразитарных систем, например, способствуя принципиальным изменениям численности хозяев паразитов в биоценозах.Эти колебания оказываются настолько существенными, что реально могут приводить к разбалансированности эволюционно сложившихсявзаимоотношений паразитов со своими хозяевами.