

Генетический код. Механизмы трансляции.
ГЕНЕТИЧЕСКИЙ КОД - это зашифровка последовательности аминокислот путем последовательности чередования нуклеотидов.
Доказательство прямого соответствия между генами и белками и раскрытие химической природы генов выдвинули на первый план вопрос о том, каким образом последовательность нуклеотидов в ДНК может программировать последовательность аминокислот в полипетидной цепи.
Поскольку и генная ДНК, и белки состоят из линейных, неразветвленных цепей, следовательно предположили, а затем и доказали, что трансляция - это последовательный процесс, в ходе которого строящийся белок наращивается по одной аминокислоте в порядке, соответствующем порядку расположения нуклеотидных оснований в гене.
Свойства генетического кода:
триплетность,
вырожденость,
универсальность,
последовательность,
неперекрываемость,
коллинеарность.
Триплетность.
Поскольку в состав белка входит 20 различных аминокислот, а в состав нуклеиновой кислоты - только 4 разных нуклеотида (А, Г, Ц, Т), последовательность нуклеотидов однозначно определяющая каждую аминокислоту, должна содержать не менее 3-х оснований, т.к. если 2 триплета кодируют 1 аминокислоту 42=16, если 43=64, если 44=256.
Сошлись на трех нуклеотидах, назвали группу из них , определяющую положение 1 аминокислоты - триплет или кодон. Долгое время эту гипотезу не удавалось проверить, первые данные подтверждающие это были опубликованы в 1961 г Криком. Он произвел тонкий генетический анализ сегмента в одном из генов фага Т4. Взаимодействие красителя акридина с ДНК бактериофага приводит к структурным изменениям в различных участках молекулы ДНК; каждое из этих изменений таково, что в ходе репликации и транскрипции все происходит так, как будто в цепь включено одно лишнее основание "вставка" или, наоборот, одно основание удалено из цепи "делеция". При многократной обработке акридитом можно получить много таких вставок (+) или "делеций" (-) по одной или по нескольку в различных сочетаниях.
ДИКИЙ ТИП
АЦТ. АЦ[Т]. АЦТ. АЦТ. АЦТ. АЦТ. АЦТ. АЦТ. и т.д.
(-)
акридин (делеция) 1-ая
МУТАНТНЫЙ ТИП
АЦТ. АЦА. Ц[Т]А. ЦТА. ЦТА. ЦТА. ЦТА. ЦТА. и т.д.
(-)
акридин (делеция) 2-ая
МУТАНТНЫЙ ТИП
АЦТ. АЦА. ЦАЦ. [Т]АЦ. ТАЦ. ТАЦ. ТАЦ. ТАЦ. и т.д. (-)
(-)
акридин (делеция) 3-ая
ДИКИЙ ТИП
АЦТ. АЦА. ЦАЦ. АЦТ. АЦТ. АЦТ. АЦТ. АЦТ. АЦТ.
измененная последовательность То же самое происходит и при вставках - при одной или 2-х модификациях одного знака + или - фаг ведет себя как мутант , однако 3 вставки или 3 делеции, а также сочетание 1 вставки и одной делеции приводит к восстановлению свойств фага. Такой результат при 3 близкородственных вставках или делециях указывал на то, что код должен состоять из 3 букв, либо из числа кратного трем, т.к. еще не было известно какому числу нуклеотидрв соответствует одна вставка или одна делеция.
В 1964 г. Ниренберг и Ледер разработали простой метод, позволивший прямо доказать 3-х буквенную структуру кода. Метод состоит в фильтровании на нитроцеллюлозном диске смеси, состоящей из рибосом, синтетических олигонуклеотидов различной длины от 2до 5-10, они использовались в качестве мРНК и различных аминоацил-т-РНК, каждая из которых несет свою аминокислоту (20 аминокислот - соответствует 20 различным тРНК). Комплекс олигонуклеотид-рибосома-аминоацил-тРНК задерживается на фильтре, а свободно? проходят через него. С помощью олигонуклеотидов, содержащих различное число остатков уридиловой кислоты У-У - ди,У-У-У - три, У-У-У-У -тетра, У-У-У-У-У -пента, было покозано, что связывается только фен-тРНК и происходит это только в том случае, если олиго-У содержит не менее трех оснований. Ниренберг и его сотрудники синтезировали все 64 возможных триплета и с помощью описанного метода не только подтвердии 3-х буквенную структуру кода для всех аминокислот, но и определили состав различных триплетов, соответствующих каждой аминокислоте.
Корниа и сотрудники (1964) готовили искусственно полирибонуклеотиды и, используя их как матрицу, синтезировали полипептидную цепь, где потом устанавливали количество и качество аминокислот.
ЛИЗ ГЛУ
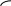
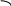
ААГААГААГ.............
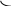
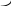
АРГ
В зависимости с чего начинали считывание получали гомопептид полиЛИЗ, ГЛУ или АРГ.
Если брали матрицы из 2-х нуклеотидов
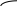
ЦУЦУЦУЦУЦУЦУЦУЦУЦУЦУ и т.д.
если бы из 4-х - гомопептид, то получалась цепь, состоящая из 2-х аминокислот