

7. Биосинтез и окисление кетоновых тел, биологическая роль этих процессов. Нарушения обмена ацетоновых тел при голодании, сахарном диабете.
Соединения ацетоуксусные и р-гидроксимасляные кислоты поступают в кровь, а затем идут в клетки тканей, но для этих молекул диффузионного барьера не существует, поэтому они служат эффективным энергетическим топливом. Эти соединения получили название - ацетоновые тела. К кетоновым телам относят три соединения близкой структуры – ацетоацетат, 3-гидроксибутират и ацетон.
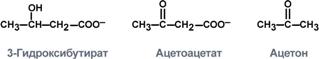
Строение кетоновых тел
Стимулом для образования кетоновых тел служит поступление большого количества жирных кислот в печень.
Синтез ацетоацетата происходит только в митохондриях печени, далее он либо восстанавливается до 3-гидроксибутирата, либо спонтанно декарбоксилируется до ацетона. Далее все три соединения поступают в кровь и разносятся по тканям. Ацетон, как летучее вещество, легко удаляется с выдыхаемым воздухом и потом. Все кетоновые тела могут выделяться с мочой.
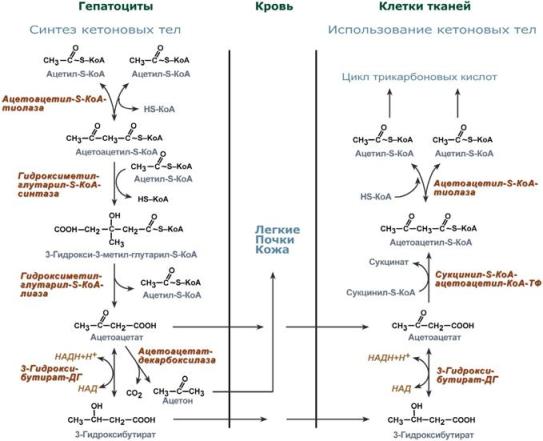
Реакции синтеза и утилизации кетоновых тел
Используются кетоновые тела клетками всех тканей, кроме печени и эритроцитов. Особенно активно, даже в норме, они потребляются миокардом и корковым слоем надпочечников.
Реакции утилизации кетоновых тел примерно совпадают с обратным направлением реакций синтеза. В цитозоле 3-гидроксибутират окисляется, образующийся ацетоацетат проникает в митохондрии, активируется за счет сукцинил-SКоА и превращается в ацетил-SКоА, который сгорает в ЦТК.
При состояниях, активирующих липолиз в жировой ткани, не менее 30% образованных жирных кислот задерживаются печенью. К таким состояниям относится голодание, сахарный диабет I типа, длительные физические нагрузки. Так как синтез ТАГ в этих условиях невозможен, то жирные кислоты из цитозоля попадают в митохондрии и окисляются с образованием кетонов. Кроме отмеченных ситуаций, количество кетоновых тел в крови возрастает при алкогольном отравлении и потреблении жирной пищи.
8. Биосинтез жирных кислот в клетках эукариот: последовательность реакций, биологическая роль. Биосинтез жиров в печени и жировой ткани. Зависимость скорости биосинтеза от режима питания и состава пищи.
Биосинтез жирных кислот наиболее активно происходит в цитозоле клеток печени, кишечника, жировой ткани в состоянии покоя или после еды.
Условно можно выделить 4 этапа биосинтеза:
1. Образование ацетил-SКоА из глюкозы, других моносахаров или кетогенных аминокислот.
2. Перенос ацетил-SКоА из митохондрий в цитозоль:
Поступающий из митохондрий цитрат в цитозоле расщепляется АТФ-цитрат-лиазой до оксалоацетата и ацетил-SКоА.
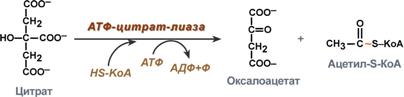
Образование ацетил-SКоА из лимонной кислоты
3. Образование малонил-SКоА из ацетил-SКоА.
Карбоксилирование ацетил-SКоА катализируется ацетил-SКоА-карбоксилазой, мульферментным комплексом из трех ферментов.
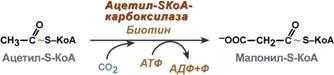
Образование малонил-SКоА из ацетил-SКоА
4. Синтез пальмитиновой кислоты.
Осуществляется мультиферментным комплексом "синтаза жирных кислот" (синоним пальмитатсинтаза) в состав которого входит 6 ферментов и ацил-переносящий белок (АПБ).
Ацил-переносящий белок включает производное пантотеновой кислоты – 6-фосфопантетеин (ФП), имеющий HS-группу, подобно HS-КоА. Один их ферментов комплекса, 3-кетоацил-синтаза, также имеет HS-группу в составе цистеина. Взаимодействие этих групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой кислоты. Для реакций синтеза необходим НАДФН.
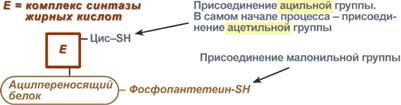
Активные группы синтазы жирных кислот
В первых двух реакциях последовательно присоединяются малонил-SКоА к фосфопантетеину ацил-переносящего белка и ацетил-SКоА к цистеину 3-кетоацилсинтазы.
3-Кетоацилсинтаза катализирует третью реакцию – перенос ацетильной группы на С2 малонила с отщеплением карбоксильной группы.
Далее кетогруппа в реакциях восстановления (3-кетоацил-редуктаза), дегидратации (дегидратаза) и опять восстановления (еноил-редуктаза) превращается в метиленовую с образованием насыщенного ацила, связанного с фосфопантетеином.
Ацилтрансфераза переносит полученный ацил на цистеин 3-кетоацил-синтазы, к фосфопантетеину присоединяется малонил-SКоА и цикл повторяется 7 раз до образования остатка пальмитиновой кислоты. После этого пальмитиновая кислота отщепляется шестым ферментом комплекса тиоэстеразой.
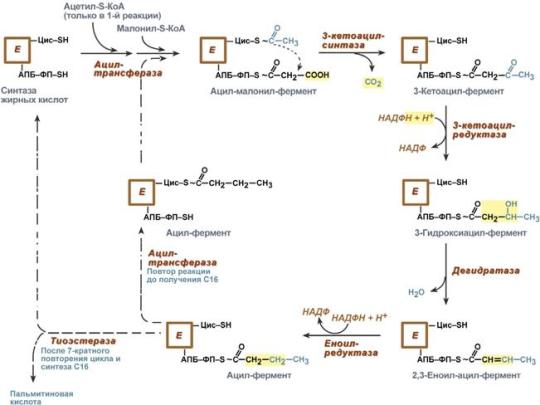
Реакции синтеза жирных кислот
9.Транспортные липопротеиды крови: особенности строения, состава, функций липопротеидов разных классов. Их роль в обмене жиров и холестерина. Пределы изменения концентрации жиров и холестерина в крови в норме.
Все липиды, присутствующие в крови, входят в состав сме
шанных надмолекулярных белковолипидных комплексов. Высшие
жирные кислоты связаны с альбуминами плазмы крови, прочие ли
пиды входят в состав липопротеидов плазмы крови. Любой липо
протеид плазмы крови состоит из монослойной амфифильной оболоч
ки, образованной молекулами апобелков, фосфолипидов, сфинго
липидов и свободного холестерола, и гидрофобного ядра, в сос
тав которого входят триацилглицерины и эфиры холестерола, а
также молекулы некоторых других липидов типа витамина Д или
витамина Е. Общее содержание липидов в ряду ХМ > ЛПОНП > ЛПНП >> ЛПВП постепенно снижается, тогда как содержание белков в том же ряду постепенно нарастает. Постепенно в том же ряду возрастает содержание фосфолипидов, а содержание триглицеридов понижается. Наконец, содержание холестерола в ряду ХМ Д> ЛПОНП Д> ЛПНП увеличивается, но затем при переходе к ЛПВП оно снижается. В зависимости от состава липопротеидных частиц они различаются по ряду свойств: плавучей плотности, электрофоре
тической подвижности и др., что используется при разделении
липопротеидов плазмы крови на ряд классов.
Белки, содержащиеся в липопротеидах, получили название
апобелков или апопротеинов. Известно несколько семейств или
классов этих белков: апоА, апоВ, апоС, апоД, апоЕ.
Апобелки различных семейств входят в состав липопротеидов
различных классов или в виде главных апобелков, или в виде ми
норных компонентов. Главными апобелками являются:
для ХМ-белки апоВ48; для ЛПОНП- апоВ100 и апоС; для ЛПНП белки апоВ100;для ЛПВП белки апоА..
1. Хиломикроны (ХМ) - образуются в клетках кишечника, их функция: перенос экзогенного жира из кишечника в ткани (в основном - в жировую ткань), а также - транспорт экзогенного холестерина из кишечника в печень.
2. Липопротеины Очень Низкой Плотности (ЛОНП) - образуются в печени, их роль: транспорт эндогенного жира, синтезированного в печени из углеводов, в жировую ткань.
3. Липопротеины Низкой Плотности (ЛНП) - образуются в кровеносном русле из ЛОНП через стадию образования Липопротеинов Промежуточной Плотности (ЛПП). Их роль: транспорт эндогенного холестерина в ткани.
4. Липопротеины Высокой Плотности (ЛВП) - образуются в печени, основная роль - транспорт холестерина из тканей в печень, то есть удаление холестерина из тканей, а дальше холестерин выводится с желчью.
Содержание холестерола в мембранах клеток периферических органов и тканей, в том числе и в клетках стенок сосудов, будет определяться сбалансированностью потоков. Явное преобладание в крови концентрации холестерола ЛПОНП+ЛППП+ЛПНП над содержанием холестерола в ЛПВП будет свидетельствовать о том, что в клетках периферических тканей накапливается холестерол и возникает угроза развития атеросклеротического процесса. Академиком А.Н.Климовым был предложен специальный показатель холестериновый коэффициент атерогенности, характеризующий соотношение этих потоков. Этот коэффициент рассчитывается по формуле в которой числитель представляет собой не что иное, как содержание холестерола ( ХС ) в ЛПОНП+ЛППП+ЛПНП. Значение этого коэффициента в норме не должно превышать 3,03,5. Если же его значение выше 3,5, человеку угрожает развитие атеросклероза.
10. Обмен и функции холестерола в организме. Биосинтез холестерола: последовательность реакции до образования мевалоновой кислоты, представление о дальнейших этапах синтеза, регуляция процесса. Роль липопротеидов крови в обмене холестерола.
Синтез холестерола в организме составляет примерно 0,5-0,8 г/сут, примерно 50% образуется в печени, около 15% в кишечнике. Все клетки организма способны синтезировать холестерол. Поступление с пищей составляет около 0,4 г/сут. Единственным реальным способом выведения холестерола является желчь – до 1 г/сут.
Биосинтез холестерола
Биосинтез холестерола происходит в эндоплазматическом ретикулуме. Источником всех атомов углерода в молекуле является ацетил-SКоА, поступающий сюда из митохондрий в составе цитрата, также как при синтезе жирных кислот. При биосинтезе холестерола затрачивается 18 молекул АТФ и 13 молекул НАДФН.
Образование холестерола идет более чем в 30 реакциях, которые можно сгруппировать в несколько этапов.
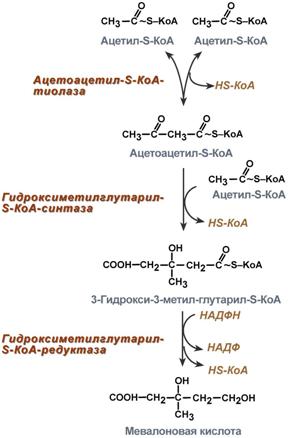
1. Синтез мевалоновой кислоты.
Первые две реакции синтеза совпадают с реакциями кетогенеза, но после синтеза 3-гидрокси-3-метилглутарил-SКоА вступает в действие фермент гидроксиметил-глутарил-SКоА-редуктаза (ГМГ-SКоА-редуктаза), образующая мевалоновую кислоту.
|
|
Реакции синтеза мевалоновой кислоты |
Схема реакций синтеза холестерола |
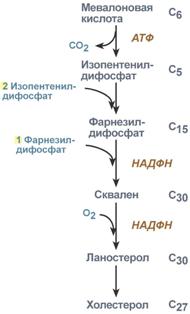
2. Синтез изопентенилдифосфата. На этом этапе три остатка фосфат присоединяются к мевалоновой кислоте, затем она декарбоксилируется и дегидрируется.
3. После объединения трех молекул изопентенилдифосфата синтезируется фарнезилдифосфат.
4. Синтез сквалена происходит при связывания двух остатков фарнезилдифосфата.
5. После сложных реакций линейный сквален циклизуется в ланостерол.
6. Удаление лишних метильных групп, восстановление и изомеризация молекулы приводит к появлению холестерола.
К периферийным тканям холестерин транспортируется хиломикроном, ЛПОНП и ЛПНП. К печени, откуда затем холестерин удаляется из организма, его транспортируют аполипротеины группы ЛПВП.