

27. Индуцированный ионный тран-т через мембрану.Затраты энерг. По формуле Борна.
Закономерности транспорта ионов через мембраны изучены в опытах с искусственными бислойными липидными мембранами (БЛМ).
Формула Борна оценивает затраты энергии для проникновения иона в неполярную фазу БЛМ:
![]()
где z – валентность иона, е – электрический заряд, r – радиус, εм – диэлектрическая проницаемость мембраны, εв - диэлектрическая проницаемость воды.
Рассчитанные по данному уравнению значения свободной энергии для перехода К+ из воды в неполярный растворитель с εм = 2 составляет большую величину (250-350 кДж/моль). Это создает барьер для прохождения гидратированного иона через гидрофобную часть липидного бислоя.
Энергия перехода иона в мембрану снижается с увеличением радиуса иона. Крупные органические молекулы проникают легче через БЛМ, чем катионы щелочных металлов.
28. Перенос ионов ионоформами. Затраты энергии для переноса комплекса «ион-ионофор». Избирательность ионных переносчиков. Электрические свойства мембраны и движение заряженных комплексов «ион-переносчик».
Величина энергетического барьера в мембране уменьшается, а следовательно, проницаемость мембраны для иона возрастает не только при увеличении его радиуса, но и при приближении значений εм к εв. На этих физических принципах основан перенос ионов ионофорами.
Ионофоры с ионом образуют комплексы большого размера – переносчики – либо формируют пору в мембране, заполненную водой, - каналы.
Перенос иона через мембрану включает стадии:
образование комплекса иона с ионофором на одной стороне мембраны,
перемещение комплекса через мембрану,
освобождение иона на другой стороне,
возвращение ионофора.
Возможны две схемы работы ионофора:
малая «карусель», когда ионофор не выходит из мембраны;
большая «карусель», когда ионофор проходит мембрану насквозь, а образование и распад комплексов протекает в неперемешиваемых слоях около мембраны.
Формула Борна описывает затраты энергии комплекса иона с радиусом r с переносчиком радиуса b c диэлектрической проницаемостью мембраны εм:
![]()
где εк эффективная диэлектрическая проницаемость внутренней сферы комплекса.
Энергия комплекса ион-переносчик значительно меньше энергии иона. Для заряженного комплекса радиусом 1 нм свободная энергия перехода из воды в мембрану составляет около 15 кДж/моль в отличие от соответствующих энергии дегидратации свободных ионов с радиусом 0,1-0,15 нм.
Ионными переносчиками являются макроциклические антибиотики, синтетические макроциклические комплексоны. Некоторые из них – нейтральные ионофоры – не содержат ионизируемых групп. Комплексоны, образуемые макроциклическими соединениями с катионами щелочных металлов, несут положительный заряд. Перенос иона происходит в виде заряженного комплекса. Катион входит во внутреннюю полость молекулы ионофора, структура стабилизируется за счет взаимодействия иона с 6-8 полярными группировками (эфирные, амидные или сложноэфирные). Ион металла экранирован от взаимодействия с растворителем.
Высокая избирательность валомицитина к образованию комплекса с К+ достигается хорошим стерическим соответствием К+ и внутренней полости валомицитина, которую образуют СО-группы. К+ полностью дегидратируется, а карбонильные (СО-) диполи заменяют гидратную оболочку (рис.5.2).
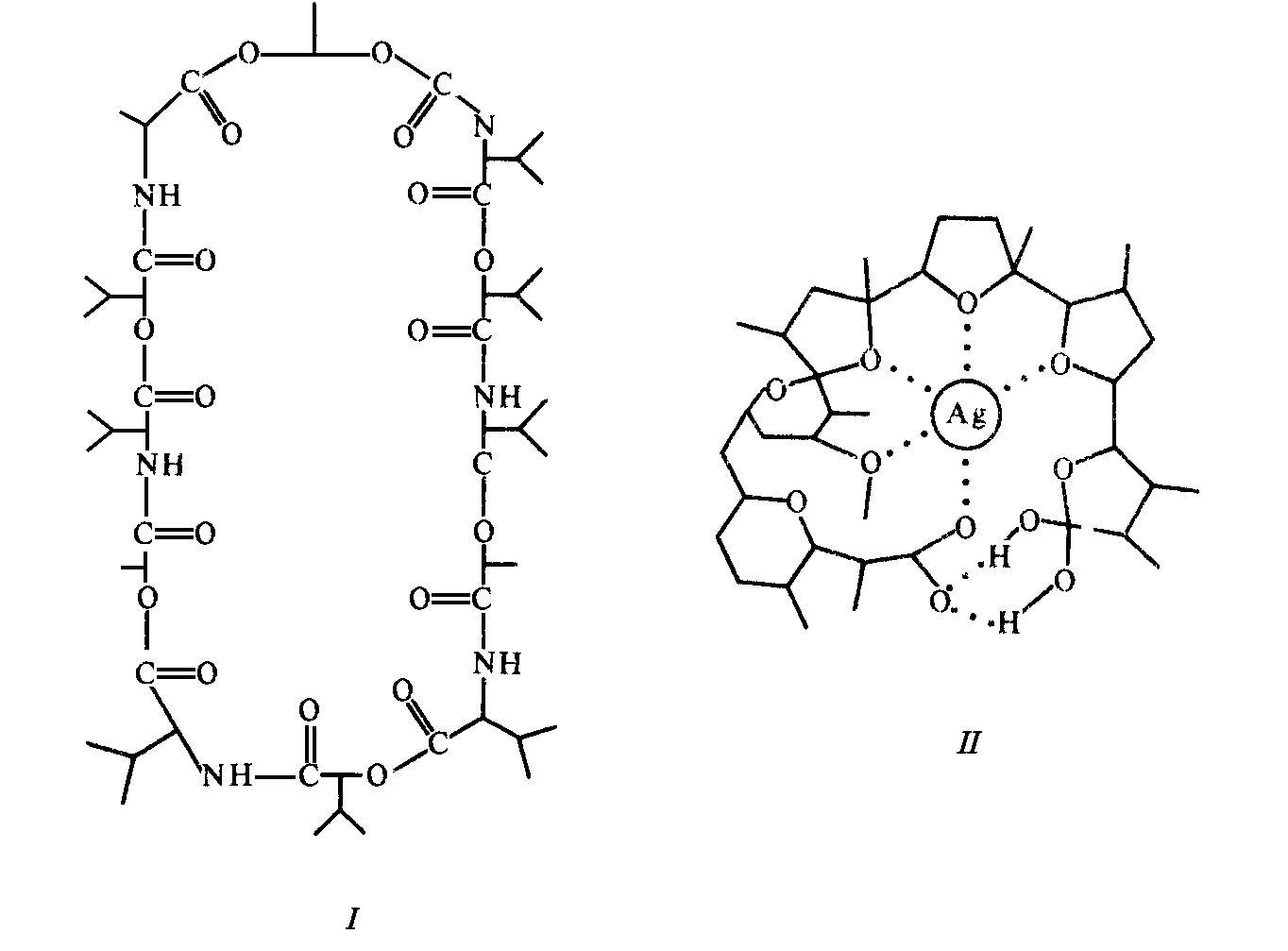
Рис. 5.2. Строение некоторых ионофоров: I — молекула валиномицина, II — система металл-кислородных и водородных связей в серебряной соли нигерицина (Рубин, 2004).
Na+ может поместиться полости валомицитина, но вследствие меньшего радиуса не взаимодействует с кислородом карбонильных групп. Энергия связывания его с водой выше энергии связывания с переносчиком.
Валомицитин обладает хорошей селективностью, переносит порядка 104 ионов/с.
Некоторые ионофоры, например нигерицин и гризориксин имеют нециклическое строение, но связывают ионы, образуя комплексную молекулу в свернутой, псевдоциклической конформации (рис. 5.2). Будучи слабыми кислотами нигерицин и его аналоги переносят два типа ионов, а при наличии градиента индуцируют обменный транспорт (обмен Н+ на К+).
Молекулы ионофора могут образовать комплекс, имеющий водную пору – канал.
Каналообразующие соединения - грамицидин А, аламетицин, амфотерицин, моназомицин и полиеновые антибиотики – обладают сродством к водной и органической фазам, поэтому образуют водную пору и сорбируются на мембране.
Полярные или заряженные группы молекулы закреплены на одной из сторон мембраны. Рассмотрим каналы, образованные грамицидином А.
Пептидная часть молекулы образована 15-ю гидрофобными аминокислотами, у которых чередуются L- и D-конфигурации. На конце молекулы формильная группа, на другом этаноламин. Каждый горизонтальный ряд аминокислот соответствует примерно одному повороту спирали, диагональные линии — пептидным связям, соединяющим отдельные витки спирали (рис. 5.3).
В мембране молекула сворачивается в спиралевидную структуру – полупору, формируя полый цилиндр длиной 3 нм и диаметром 0,5-0,8 нм. Непроводящее состояние молекулы не имеет внутренней полости. Равновесие между формами смещается под действием внешнего электрического поля. Грамицидин в субмолекулярных концентрациях повышает проводимость БЛМ в 106-108 раз.