

17 Билет
Вторичное строение корня характерно только для двудольных покрытосеменных и голосеменных растений, т. е. для растений, имеющих вторичные меристемы — камбий и феллоген. В формировании вторичного строения участвует только центральный цилиндр, а первичная кора и ризодерма отмирают и слущиваются. Центральный же цилиндр разрастается в ширину за счет работы камбия, который закладывается из прокамбия, оставшегося между ксилемой и флоэмой радиального проводящего пучка. Снаружи корень покрывается пробкой, образовавшейся из феллогена перициклического происхождения. За исключением первичной ксилемы проводящего пучка, расположенной в центре органа, все ткани корня вторичного строения имеют вторичное происхождение.
18.Специализация и метаморфоз корней растений
Некоторые корни выполняют особые функции, в связи с чем меняется и их строение.
Очень часто молодые корневые окончания образуют симбиоз с гифами почвенных грибов , называемый микоризой (дословно "грибокорень"). Чаще всего микоризой становится коровая часть корня в зоне всасывания. Высшее растение и гриб извлекают из такого симбиоза взаимную пользу. Во многих случаях гифы грибов функционально заменяют корневые волоски. Большая часть многолетних растений имеет микоризу. Предполагается, что микориза является одним из факторов, способствовавших прогрессу цветковых растений.У эпифитных тропических орхидей и некоторых других растений корни покрыты многослойной эпиблемой , называемой веламеном . Веламен выполняет иногда фотосинтезирующую функцию, а позже может участвовать в поглощении атмосферной влаги, образуя мертвый губчатый гигроскопичный покров корня.
На корнях бобовых и ряда видов других семейств возникают особые образования - клубеньки , в которых поселяются бактерии из рода Rhizobium или реже. Эти микроорганизмы способны фиксировать атмосферный молекулярный азот, переводя его в связанное состояние. Часть азотистых соединений, образовавшихся таким путем, усваивает высшее растение-хозяин. С другой стороны, бактерии используют вещества, находящиеся в корнях высшего растения. Описанный симбиоз крайне важен для сельского хозяйства, так как благодаря ему почва обогащается азотистыми веществами.
Очень широко распространены так называемые втягивающие, или контрактильные, корни, обычные у многих луковичных и корневищных растений. Укорачиваясь у основания, такие корни способны втягивать луковицы или корневища в почву на оптимальную для их сохранения глубину в период летней засухи или зимних морозов.
У растений, живущих на бедных кислородом почвах ( таксодиум, или болотный кипарис , растения затопляемых морских побережий, образующих мангровые заросли), имеются дыхательные корни - пневматофоры . Некоторые тропические деревья развивают от оснований стволов досковидные корни, служащие своего рода подпорками.Очень широко распространены запасающие корни. Возникновение запасающих корней нередко приводит к резкомувидоизменению всей корневой системы. Стержневая корневая система, в которой запасающим стал главный корень, нередко превращается в так называемый "корнеплод" ( морковь , петрушка , сельдерей ). Однако у ряда "корнеплодов" большая их часть образована основанием стебля и разросшимся гипокотилем , а собственно корень представляет собой лишь самую нижнюю часть "корнеплода", несущую боковые корни. "Корнеклубни" возникают при видоизменениях ветвистой и мочковатой корневых систем ( георгина , некоторые виды орхидных ).
Корнеплод.
Каждый корнеплод по вертикали делится на три части (зоны): головку, шейку и собственно корень. Головка - верхняя часть корнеплода, несущая на себе листья и почки. Шейка -расположена ниже головки, не имеет ни листьев, ни боковых корней. Собственно корень - нижняя часть корнеплода, на ней располагаются боковые корни, это позволяет отличить ее от шейки. По наличию питательных веществ шейка и собственно корень - полноценные части корнеплода.
Снаружи корнеплод покрыт покровной тканью-пробкой, внутрь от которой располагаются лубяная (флоэма) и древесная (ксилема) части корнеплода, состоящие главным образом
из запасной паренхимы, богатой питательными веществами. Внутреннее строение отдельных корнеплодов неодинаковое; различают корнеплоды типа моркови, редиса и свеклы
ВОПРОС № 19. АНАТОМИЧЕСКОЕ СТРОЕНИЕ ЛИСТА. СВЯЗЬ ВНУТРЕННЕГО СТРОЕНИЯ ЛИСТА С ЕГО ФУНКЦИЯМИ.Лист – орган второго порядка, занимающий боковое положение на стебле (оси побега) и выполняющий функции ! фотосинтеза, транспирации(регулирование испарения воды) и газообмена! Лист как правило состоит из основания, пластинки и черешка. Анат строение листа изучается на примере листовой пластинки, у кот имеется верхняя(брюшная,адаксиальная) и нижняя(спинная,абаксиальная) стороны.У листа только первичные ткани. Снаружи лист покрыт прозрачной эпидермой(покровная ткань молодых стеблей и листьев). Между верхней и нижней эпидермой располагается комплекс тканей =мезофилл, кот. вкл. Столбчатую ткань или палисадную паренхиму (наиболее развиты хлоропласты,вытянутые кл. ценр часть занимает вакуоль) и губчатую паренхиму (также сод-т хлорофилл,ц.часть занята вакуолью, форма клеток округлая с рыхлым расположением) В эпидерме клетки располагаются плотно, антиклинальные (перпенидк-но поверхности) стенки, извитые, К эпидерме относятся основные эпидермальные клетки, трихомы(отдельн клетки) и замыкающие кл устьиц. Наружные стенки осн-х эпидермальных клеток покрыты кутикулой для водоотталкивающих свойств. Устьица состоят из замыкающих клеток, устьичной щели и подустьичной полости. Устьица есть как на ниж так и на верхней поверхности листа. Часть устьиц всегда открыта, благодаря этому вода поднимается по растению.Проводящие пучки в листе всегда закрытые! В проводящ пучке листа ксилема всегда обращена к верхней стороне листа, флоэма к нижней, так как проводящ системы листа и стебля соединены.В жилках есть механическая ткань, склеренхима,окружающая проводящие ткани для защиты пучка от сдавливания. Волокон склеренхимы в листе нет,только вдоль жилок. А в самой ткани листа есть идиобласты склеренхимы (склереиды - механич клетки в) для укрепления ткани листа. В клетках листа часто встречаются кристаллы минеральных солей (назначение – особождение от конечных продуктов обмена веществ при листопаде). Дополнительные функции листа запасание воды (субкуленты - алоэ) и сахаров (сочные чешуи луковиц – это ниж часть листа), размножение.
ВОПРОС№20 РАЗНООБРАЗИЕ ЛИСТЬЕВ. ФОРМАЦИИ. ГЕТЕРОФИЛИЯ И АНИЗОФИЛИЯ. ЛИСТОВАЯ МОЗАИКА.
Главная часть ассимилирующего листа – пластинка. Различают листья с одной пластинкой – простые, и сложные с несколькими пластинками. Общую ось сложного листа, несущую листочки, называют рахисом. В зависимости от расположения листочков на рахисе различают перисто и пальчато- сложные листья, и тройчатый с тремя пластинками (клевер).Простые листья могут быть перисто- и пальчато-лопастные, перисто- и пальчато- раздельные, перисто и пальчато- рассеченные.
Листья не одинаковы не только у разных растений но и у одного растения. Формации листьев : низовой формации (чешуевидные), срединные (собственно зеленые листья) и верховые или прицветные (верховой формации) – образуются в области соцветия и служат кроющими листьями отдельных цветков соцветия, защищая их в почке.
Гетерофиллия (гетеро-разный, филлон-лист) разнолистность, -различие в форме, размерах и структуре листьев на одном и том же растении, в узком смысле – различия между листьями срединной формации в пределах побега. Различают возрастную и экологическую гетерофиллию. (водяной лютик)
Анизофиллия (анизос-неравный, филлон – лист) – различие в форме размерах и структуре ассимилирующих листьев на одном и том же узле побега (при супротивном и мутовчатом листорасположении). Причины – действие силы тяжести и неравномерное освещение верхней и нижней стороны побега, но чаще наследственно закреплена. (конский каштан)
Листовая мозаика - расположение листьев в горизонтальной плоскости , при к-ром обеспечивается наименьшее затенение их друг другом. Листовая мозаика образуется благодаря фототропизму листьев, гетерофиллии и анизофиллии, неравномерному росту черешков. Ярко выражена на плагиотропных (плагиотропизм – способность боковых органов растений принимать положение под опред углом к осевому органу в зависимости от тяжести света идр) побегах, вертикально растущих розеточных побегах (липа клен плющ герань подорожник)
ВОПРОС№21 АНАТОМИЧЕСКОЕ СТРОЕНИЕ СТЕБЛЯ ОДНОДОЛЬНЫХ ТРАВЯНИСТЫХ РАСТЕНИЙ.
Для стеблей однодольных растений характерно отсутствие вторичного утолщения. Проводящие пучки однодольных закрытые, то есть не имеют камбия! И поэтому все элементы пучков первичного происхождения. Они возникли из клеток прокамбия путем дифференциации. Все остальные части стебля также образованы первичными тканями. Многочисленные тяжи прокамбия в конусе нарастания , а значит и проводящие пучки проходят не по вертикали вдоль стебля, примерно на одинаковом расстоянии от центра стебля, как у двудольных, а на своем протяжении в междоузлии образуют дуги, отходящие от периферии к центру и загибающиеся вновь к периферии с тем, чтобы в узлах стебля проследовать в листья. Таким образом все проводящие пучки стебля однодольных представляют собой листовые следы или следы пазушных почек. Вследствие этого на поперечных срезах стеблей большинства однодольных пучки расположены не в виде кольца, а разбросаны по всей толще центрального цилиндра так что в ее основной паренхиме нельзя выделить сердцевину и сердцевинные лучи. Сердцевина образуется только в стеблях представителей немногих семейств. В стеблях однодольных нельзя выделить первичную кору – центральный цилиндр начинается сразу под эпидермой. Для изучения типичного строения стебля однодольных удобны стебли растений семейств лилейных и ирисовых, кукурузы, ржи пшеницы и др злаков.Строение стебля ириса: снаружи эпидерма и толстая кутикула. Под эпидермой первичная кора, в состав которой входит хлоренхима и эндодерма. Дальше располагается центральный цилиндр начинающийся красным кольцом склеренхимы. Слой склеренхимы представляет собой многорядный перицикл, внутрь от которого среди крупных клеток основной паренхимы расположены закрытые коллатеральные пучки (состоят из механической обкладки пучка, склеренхимы, флоэмы и ксилемы). Проводящие пучки расположены в беспорядке, но на периферии их больше и мелких , в центре стебля – меньше, но они крупнее.
22анатомическое строение стебля двудольных травянистых растений.Стебель представляет собой ось побега, несущая листья и почки. Основные функции стебля - опорная и проводящая. Стебель осуществляет связь между корнями и листьями. Кроме того, в стебле нередко откладываются запасные питательные вещества. Иногда стебель - ассимилирующий орган. анатомическая структура стебля, сохраняющаяся у однодольных в течение всей жизни. У двудольных и голосеменных довольно быстро нарушается в результате разного рода вторичных изменений и в итоге формируется так называемое вторичное строение стебля. В результате деятельности прокамбия и остальной первичной меристемы конуса нарастания образуется первичное строение стебля. В первичном стебле обычно различают первичную кору и стелу (центральный цилиндр). В отличие от корня первичная кора снаружи покрыта эпидермой. Сердцевина расположена в центре стебля и состоит преимущественно из паренхимы. Сердцевина многих растений частично разрушается, и тогда стебель становится полым. В стебле сердцевина сообщается с первичной корой при помощи паренхимной ткани, расположенной радиальными рядами и получившей название сердцевинных лучей. При любом типе вторичных изменений в центральном цилиндре первичная ксилема <оттесняется> к центру и остатки ее располагаются на границе с сердцевиной. Напротив, первичная флоэма оттесняется нарастающей вторичной флоэмой к периферии и в дальнейшем становится малозаметной.
23. анатомическое строение стебля двудольных древесных растений. У древесных и кустарниковых двудольных, а также у хвойных вторичные утолщения могут продолжаться многие годы. В итоге в стебле выделяют три основные части: кору, древесину и сердцевину. Граница коры и древесины проходит по камбию. Кора многолетнего стебля древесного растения включает перидерму, остатки первичной коры, группы механических элементов различного происхождения, располагающихся на границе остатков первичной коры и флоэмы, и всю массу флоэмы (вторичную флоэму - луб и остатки первичной). У ряда древесных растений с возрастом на смену перидерме формируется корка (ретидом). Луб дифференцирован на мягкий луб, состоящий из проводящих и паренхимных элементов. Совокупность механических элементов вторичной флоэмы получила название твердого луба. Годичные кольца наиболее характерны для многолетних древесных растений умеренных широт, когда периоды интенсивного весенне-летнего роста камбия чередуются с временами осенне-зимнего покоя. Каждое кольцо состоит из двух частей — светлой и тёмной. У хвойных деревьев годичные кольца хорошо видны так как их поздняя древесина принимает тёмный цвет. По количеству колец на спиле судят о возрасте дерева, а также о темпах его роста, в зависимости от того на какой высоте был сделал поперечный спил.
24разнообразие побегов по функциям, длине междоузлий, направлению роста, положению в пространстве. Смена направлений роста одного и того же побега.Побе́г — один из основных вегетативных органов высших растений, состоящий из стебля с расположенными на нём листьями и почками. различают побеги вегетативные, вегетативно-генеративные и генеративные. Вегетативные (не видоизменённые) побеги, состоящие из стебля, листьев и почек и вегетативно-генеративные (частично видоизменённые), состоящие дополнительно из цветка или соцветия, выполняют функции воздушного питания и обеспечивают синтез и неорганические. В генеративных (полностью видоизменённых) побегах фотосинтез чаще всего не происходит, зато там образуются спорангии, задача которых сводится к обеспечению размножения растения (к таким побегам относится и цветок).Побег, на котором образуются цветки, называется цветоносным побегом, или цветоносом. .длина междоузлий- Тот участок стебля, от которого отходит лист и почка в его пазухе, называют узлом. Участок стебля между соседними узлами – это междоузлие. Таким образом, вегетативный побег состоит из частей, повторяющихся по его длине: узлов с листьями и пазушными почками и междоузлий. направлению роста- Первый побег проростка обычно растет вертикально вверх. Он прямостоячий. Но в дальнейшем, когда из почек образуются и другие побеги, они, как и корни в корневой системе, могут оказаться разными по направлению роста. У дерева ствол обычно прямостоячий. То же можно сказать об отдельных стволиках кустарников. У многих трав, однолетних и многолетних, побеги тоже прямостоячие, например у ржи, подсолнечника, ромашки, колокольчика. пиона, гладиолуса. У трав, кроме прямостоячих, различают приподнимающиеся, лежачие, ползучие побеги. Приподнимающиеся побеги имеют изгиб в нижней части: сначала они растут горизонтально, потом вертикально. Прямостоящие побеги- Растут вертикально вверх и не нуждаются в опоре. Ползучие побеги- Стелются по земле и укореняются в почве при помощи придаточных корней. Приподнимающиеся побеги- Основания побегов занимают горизонтальное положение, а верхняя часть – вертикальное. Цепляющиеся побеги- Поднимаются вверх, прикрепляясь к опоре усиками, или корнями-зацепками. Вьющиеся побеги- Выносят листья к свету, обвиваясь вокруг прямостячих стеблей или искуственных опор.
25 метаморфозы надземных побегов.Метаморфоз — превращение одного органа в другой с полной сменой формы и функции,довольно распространенным типом видоизменения стебля является колючка. Колючка представляет собою укороченный побег с заостренной верхушкой. Она возникает обычно в пазухе листа и на ней часто можно видеть листья и цветки, как например у Коллеции колючейшей. У Боярышника однопестнчного колючки весною развиваются как пазушные побеги с нормальными листьями и с верхушечной почкой; если побеги в этом состоянии срезать и поместить в воду, они развиваются в нормальные побеги. И а растении же эти побеги вскоре одревесневают, листья недоразвиваются и опадают, оставляя колючку голой.У ряда растений боковые побеги видоизменены в усики служащие для прикрепления к опоре. Видоизмененные в усики побеги особенно характерны для тыквенных, а также для винограда.Своеобрааными видоизменениями ползучего побега являются усы Они представляют собою плагнотроппые побеги, возникающие в пазухах низовых листьев усы обычно тонкие стелющиеся по земле стебли, с более или менее удлиненными междоузлиями, концы которых, укореняясь, образуют розетку листьев. Из последней возникают новые плети. усы недолговечны (существуют не более одного вегетационного периода) и с отмиранием междоузлий розетки становятся самостоятельными особями.Усами обладают Земляника лесная Лапчатки гусиная.Филлодии— листоподобное расширение черешка, которое выполняет функции листовой пластинки частично или полностью редуцированной.Филлокладии— видоизменённые побеги растений, у которых стебли приобретают листовидную форму и выполняют функцию фотосинтеза, а листья редуцированы и представлены чешуйками, расположенными по краям или на поверхности филлокладия. В пазухах этих чешуевидных листьев развиваются соцветия или одиночные цветки.Кладодий видоизмененный побег растений с уплощенным, листовидной формы стеблем, выполняющий функции листа. Листья на К. сильно редуцированы, превращены в колючки или рано опадают.
26 метаморфоз подземных побегов Метаморфоз — превращение одного органа в другой с полной сменой формы и функции, Подземные побеги встречаются у многолетних растений с зелеными надземными побегами. Они выполняют функцию запасающих органов, в которых растение откладывает в летний период запасные вещества. Подземные побеги нужны также для вегетативного возобновления. Весной из них вырастают надземные зеленые побеги, питающиеся запасом питательных веществ из подземных. Среди видоизменений подземных побегов можно назвать корневища, клубни и луковицы. Корневище – подземный продолговатый побег с укороченными междоузлиями. Обычно они утолщенные, иногда мясистые, расположены в земле горизонтально или наклонно. Корневище имеет на одном конце верхушечную почку, этим концом оно растет; противоположный конец его постепенно отмирает. В узлах побега находятся мелкие, чешуйчатые листочки, а в их пазухах – боковые почки. Из боковых и верхушечной почек вырастают надземные побеги. На нижней стороне корневища образуются мочковатые придаточные корни. Длинные, тонкие, разветвленные корневища, ненакапливающие питательные вещества, называются подземными ползучими побегами. Они служат для вегетативного возобновления. Клубень является сильно утолщенным, коротким, вздутым подземным побегом, накапливающим запасные питательные вещества, На его поверхности в углублениях находятся почки, одиночные или собранные по несколько штук в пазухах чешуйчатых быстро опадающих листочков Из этих почек весной вырастают зеленые надземные побеги. Некоторые растения имеют одновременно два клубня, один старый, другой новый, последнего года.
Луковица состоит из очень укороченного, конусовидного стебля, образующего, так называемое, донце, а также окружающих его незеленых чешуйчатых или мясистых листьев, в которых накапливаются запасные вещества. Листья плотно прилегают друг к другу и образуют большую луковицу, покрытую внешними, сухими, защитными чешуями. Из луковицы весной вырастает зеленый, облиственный, цветоносный побег. Новые луковички образуются в мясистых пазухах старых чешуек и служат для вегетативного возобновления. Такие подземные побеги встречаются у растений однодольных, а также с коротким периодом вегетации. Следующим типом видоизменения подземного побеги являются столоны Так называется подземный, быстро растущий побег, несущий на конце почку, клубень или луковицу. в отличие от корневищ столоны, как правило, не долговечны. Кроме того, они обладают ограниченным ростом, который прекращается с формированием ночки, клубня или луковицы.особую форму видоизмененного стебля представляет клубнелуковица.она формируется из укороченного утолщенного стебля несущего на верхушке или у основания почку из которой развиваются листья и цветки из нижней части образуются адвентипные корни.клубнелуковица покрыта остатками оснований прошлогодних листьев располагающихся по спирали в пазухах которых возникают новые клубнелуковицы или побеги.клубнелуковицы свойственны многолетним растениям однако сама клубнелуковица не является многолетней,из нее развиваются вегетативные и генеративные органы расходующие весь запас питательных вещ-в накопленных в ней после чего она отмирает
27.Соцветия, классификация соцветий.Соцветия – побег или система побегов несущих цветки.(присущи большинству цветковых растений) Характеризуются тем, что цветки распускаются на осях снизу вверх.биологическое значение - повышение вероятности перекрестного опыления.В зависимости от степени ветвления соцветия делят на:Простые - на главной оси располагаются одиночные цветки, ветвление не превышает 2 порядков;Сложные - на главной оси располагаются боковые оси, т.е. ветвление достигает 3-4 порядков.Нарастание осей может быть:Моноподиальным (способ нарастания при котором ось формируется за счёт деятельности верхушечной почки),Симподиальным (способ нарастания при котором каждая ось формируется за счёт деятельности верхних боковых почек) Тирс сложное соцветие в к-ром главная ось нарастает моноподиально а боковые симподиально.По наличию и характеру прицветных листьев:Фрондозное прицветкини сходны с ассимиляционнными листьями,Брактеозное кроющие листья цветков представлены прицветниками,Голое соцветие-прицветники редуцированы.Простые моноподиальные соцветия:Простые соцветия с удлиненной осью:Кисть. характеризуются удлинённой главной осью и цветками на хорошо выраженных цветоножках более или менее одинаковой длины. Различают несколько видов: фроидозные (фиалка трехцветная), брактеозные (люпин, черемуха), голые (капуста, сурепка), открытые (гиацинт), закрытые (колокольчик),Щиток нижние цветоножки намного длиннее верхних и все цветки располагаются в одной плоскости, (калина, груша, боярышник),Колос соцветие, для которого характерна удлинённая главная ось, на которой расположены сидячие одиночные цветки или колоски из нескольких цветков (подорожник)Сережка соцветие с удитненной повислой осью на которой скученно расположены мелкие цветки. (ива, тополь),Початок соцветие с удлиненной толстой мясистой главной осью и сидячими цветками
Простые соцветия с укороченной осью:Зонтик соцветие с сильно укороченной главной осью несущей на верхушке цветки с удлененными цветоножками почти равной длины(примула, чистотел, женьшень),Головка соцветие с укороченной главной осью и сидячими на коротких цветоножках цветками (клевер),Корзинка соцветие с разросшейся плоской вогнутой или блюдцевидной главной осью на которой плотно прилегая друг к другу сидят цветки(сем.Астровые)
Сложные моноподиальные соцветия :Сложный щиток (соцветие, главная ось которого несет боковые ветвящиеся оси, заканчивающиеся простыми щитками. тысячелистник, рябина),Сложный колос соцветие, образованное несколькими простыми колосками, сидящими на одной общей цветоножке. – пшеница, рожь, ячмень,Сложный зонтик соцветие, состоящее из многих простых зонтиков (сем.Сельдерейные – морковь, петрушка),Метелка на главной оси развиваются боковые оси несущие простые сроцветия (сирень).Симподиальные соцветия – (закрытые) у них главная ось не выражена, а парциальные соцветия развиваются только в непосредственной близости от верхушечного цветка.Монохазии главная ось заканчивается цветком под ним образуется боковая ось с одним цветком.Завиток (бурачник) – все оси направлены в одну сторону.Извилина (гладиолус) – все оси направлены в разные стороны.Дихазии– соцветие, в котором подверхушечным цветком главной оси развиваются 2 побега с верхушечнымицветками Плейохазии от главной оси заканчивающейся цветком отходит несколько боковых осей Тирсы
34. Ценокарпные плоды – плоды, образованные сросшимися между собой плодолистиками. Срастание плодолистиков между собой имеет большое приспособительное значение – лучшая защита семязачатков. Плоды-ценокарпии - самая многочисленная группа плодов. Морфологические типы ценокарпиев весьма разнообразны. Главнейшие из них – ягода (многосемянный сочный плод с тонким кожистым экзокарпием и сочным мезокарпием и эндокарпием. Встречается во многих семействах: виноградные, лилейные, брусничные и др.), коробочка (сухой многосемянный плод. Встречается во многих семействах: луковые, фиалковые, колокольчиковые и др.), стрючок (сухой многосемянный вскрывающийся плод с пленчатой ложной перегородкой, представляющей собой вырост плацентарных бороздок. Характерен для крестоцветных), вислоплодник (сухой двусемянный дробный плод, распадающийся на два мерикарпия, повисающих на двураздельной колонке, возникающей из брюшных проводящих пучков плодолистиков. Характерен для большинства зонтичных и некоторых аралиевых), а также листовка (сухой многосемянный мономерный вскрывающийся по брюшному шву плод), яблоко (сочный плод растений подсемейства яблоневые), тыквина (сочный многосемянный плод растений семейства тыквенные), гесперидий (характеризуется железистым экзокарпием, губчатым мезокарпием и разросшимся эндокарпием, имеющим вид соковых мешочков. Характерен для цитрусовых), ценобий (дробный плод, образующийся из двух плодолистиков и распадающийся на четыре односемянные части. Характерен для губоцветных, бурачниковых), костянка (сочный плод с резко выраженной дифференциацией слоев околоплодника: экзокарпий – тонкий кожистый, мезокарпий – сочный, эндокарпий – одревесневший. Н-р: персик, калина, миндаль, крушина).
35. Высшие растения – это растения, тело которых дифференциировано на вегетативные органы. Эволюция большинства высших растений шла по пути приспособления к условиям наземного существования. К высшим растениям относятся отделы: риниофиты, моховидные, плауновидные, псилотовидные, хвощевидные, папоротниковидные, голосеменые и покрытосеменные. Предками высших растений были какие-то морские водоросли, у которых, в связи с переходом в наземные условия среды выработались специальные приспособления для водоснабжения, для защиты гаметангиев от высыхания и для обеспечения полового процесса. Признаки современных высших растений можно разделить на три группы: а) унаследованные от водорослевых предков в неизменном виде: макроскопические размеры и многоклеточное строение тела, одноядерность клеток, зеленая окраска и пигментный состав пластид(хлорофилл, каротин, ксантофилл), основной продукт запаса – крахмал, эндогамный половой процесс, жизненный цикл диплогаплофазный со споротической редукцией; б) признаки водорослевых предков, достигшие у высших более высокого уровня дифференциации: зеленые пластиды в виде многочисленных мелких линзовидных хлорофилловых зерен без пиреноидов, ярко выраженная относительно устойчивая морфолого-физиологическая полярность тела, строгая локализация меристем, в частности, наличие боковых меристем и вторичного утолщения, многоклеточные, иногда многослойные стенки у спорангиев и гаметангиев, инкапсулированный зародыш в начальных стадиях развития; в) вновь приобретенные признаки адаптации к наземным условиям существования: способность клеточных оболочек к кутинизации, одревесневению и опробковению, формирование высоко специализированных тканей, таких как эпидерма, ксилема, флоэма, колленхима, склеренхима, целостная проводящая система тканей – стелла, четко выраженная морфофизиологическая дифференциация тела на побег(стебель, листья, почки) и корень.
36. Высшие растения обладают гетероморфными циклами развития. Высшие растения приспособлены к существованию на суше. Совершенствование этой приспособленности прослеживается и при рассмотрении особенностей их строения, связанных с размножением. Все высшие растения оогамны. Их типичные гаметангии многоклеточны. Мужские гаметангии называются антеридиями, в них образуются мужские половые клетки (гаметы) - сперматозоиды или спермии. Женские гаметангии называются архегониями, в них образуются женские гаметы - яйцеклетки. Все приспособления выражались в возрастающем расхождении в строении гаметофита и спорофита. В жизненном цикле высших растений закрепилось, имевшее место еще у водорослей, чередование двух фаз - полового (гаметофита) и бесполого (спорофита). На гаметофите развиваются органы полового размножения, а на спорофите образуются спорангии, внутри которых, в результате редукционного (мейотического) деления, образуются споры с гаплоидным (n), набором хромосом. Гаметофит может быть обоеполым, когда на нем развиваются и антеридии, и архегонии, или однополым, тогда мужской гаметофит несет антеридии, а женский - архегонии. При слиянии мужской и женской гамет образуется диплоидная зигота с диплоидным набором хромосом. Из зиготы (она делится митотически) развивается диплоидный спорофит. Если образующиеся споры на спорофите одинаковы, то из них развивается обоеполый гаметофит. Но у большинства высших растений в спорангиях образуются споры двух типов (в разных спорангиях): в макроспорангиях образуются макроспоры, а в микроспорангиях - микроспоры. Из каждой микроспоры развивается один мужской гаметофит, а из макроспоры (мегаспоры) - один женский. Поэтому микроспоры называют мужскими спорами, а мегаспоры - женскими. Полный жизненный цикл высшего растения состоит из гаметофита (период от споры до зиготы) и спорофита (период от зиготы до образования спор). Все высшие растения имеют еще и зародыш. Из зиготы у высших растений всегда развивается зародыш (эмбрион) - зачаток новой особи - спорофита (2n). Только у цветковых зародыш формируется за счет запасных продуктов спорофазы - вторичного эндосперма и является триплоидным. Эволюция жизненного цикла высших растений шла в двух направлениях: у моховидных - по возрастанию самостоятельности гаметофита с его морфологическим расчленением, потерей самостоятельного спорофита и его упрощением (до спорогона, существующего на гаметофите), тогда как у всех остальных высших растений преобладающей фазой становится спорофит, а гаметофит уменьшается и упрощается. Таким образом, эволюция высших растений, за исключением моховидных, шла по пути редукции гаметофита и совершенствования спорофита.
37. Моховидные, или Мхи, или Настоящие мхи, или Бриофиты (лат. Bryophyta) — отдел высших растений. насчитывающий около 10 000 видов, объединённых примерно в 700 родов и 110—120 семейств. это мелкие растения, длина которых лишь изредка превышает 50 мм; исключение составляют водные мхи, некоторые из которых имеют длину более полуметра, и эпифиты, которые могут быть ещё более длинными. Моховидные, как и другиеМохообразные, отличаются от других высших растений тем, что в их жизненном цикле гаплоидный гаметофитпреобладает над диплоидным спорофитом.Многие данные указывают на то, что мхи, как и другие споровые растения, произошли от псилофитов (риниофитов) — группы древних вымерших наземных растений. При этом спорофит мхов рассматривается как конечный результат редукции разветвлённого спорофита предковых форм.Есть и другая точка зрения, согласно которой мхи, плауновидные и риниофиты имеют общее происхождение от ещё более древней группы растений.[3]Ранние палеонтологические находки датируются концом девона — началом карбона[4].У мхов нет цветков, корней и проводящей системы. Размножаются мхи спорами, которые созревают в спорангиях на спорофите. В жизненном цикле, в отличие от сосудистых растений, преобладает гаплоидный (то есть с одинарным набором непарных хромосом) гаметофит (половое поколение). Гаметофит мхов — многолетнее зелёное растение, нередко с листоподобными боковыми выростами и корнеподобными выростами (ризоидами), в то время как спорофит (или бесполая стадия жизненного цикла) короткоживущий, быстро усыхает и состоит только из ножки и коробочки, в которой созревают споры.Спорофит моховидных (носящий название спорогоний, или спорогон), имеет более простое строение, чем у других групп высших растений. Он не способен укореняться и располагается на гаметофите. Спорофит состоит, как правило, из трёх элементов:коробочка (или спорангия), в которой развиваются споры;ножка (или спорофора), на которой располагается коробочка;стопы, обеспечивающей физиологическую связь с гаметофитом.Мхи встречаются на всех континентах, в том числе в Антарктиде, нередко в экстремальных условиях обитания.Обычно мхи образуют плотные скопления в затенённых местах, нередко в непосредственной близости от воды, хотя могут встречаться и на относительно открытых, сухих участках. Среди моховидных есть виды, живущие в пресноводных водоёмах. Морских обитателей нет, хотя некоторые виды поселяются на скалах в полосе прибоя.
38. Печёночные мхи, или Печёночники, или Маршанциевые мхи (лат. Marchantiophyta, Hepatophyta) — отдел растений со слабо развитой протонемой;Печёночные мхи — мелкие и нежные мохообразные растения.Одни из них снабжены стебельками и листьями, лишенными всяких жилок и расположенными в два или три ряда; те, что находятся на стороне, обращенной к почве, чешуевидны и совершенно другой формы, чем остальные. Верхние листья, располагаясь обыкновенно в два ряда, имеют две лопасти, из которых одна маленькая принимает особую форму и пригнута книзу. Таковы листостебельные печёночницы.Другие представляют плоское или плосковатое слоевище, распростёртое на земле или даже плавающее на воде. Это слоевище обыкновенно ветвится развилисто, тёмно-зелёного цвета и несёт у некоторых, на нижней стороне, нежные чешуйки, расположенные в два ряда и соответствующие, очевидно, листьям.Мужские (антеридии) и женские (архегонии) органы этих растений построены совершенно наподобие тех же органов лиственных мхов, но расположены они часто иначе, иначе и развиваются после оплодотворения. У листоносных они, подобно лиственным мхам, занимают верхушки стебельков, у слоевцовых — или образуются особые отростки разной формы, несущие на верхушке органы оплодотворения, или же эти органы сидят на самом слоевище. После оплодотворения нижняя вздутая часть архегония, разрастаясь, не выносится, как у лиственных мхов, вверх, а пробуравливается развивающимся спороплодником и остаётся при основании его ножки в виде нежного влагалища. Самый спороплодник раскрывается створочками или зубцами на 2, 4 и больше; у многих он содержит, кроме спор, так называемые элатеры, то есть веретенчатые, извилистые клеточки, содержащие спиральное утолщение внутри; элатеры, вследствие своей гигроскопичности, действуют как пружинки, способствуя раскрыванию спор.Кроме полового размножения, печеночные мхи способны размножаться с помощью особых почек, содержащихся, как, например, у маршанции, в особых блюдцевидных вместилищах. Заросток у этих растений незначительный, а корни заменены одноклетными волосками, обильно образующимися на нижней стороне слоевища или на основании стебельков.Для маршанции, как и для других Печеночников, характерно чередование поколений с доминированием гаметофита (n) над спорофитом (2 n). Два важнейших события в жизненном цикле – мейоз и сингамия. Гаметофитное поколение начинается с мейоза, дающего гаплоидные споры. Одни из них прорастают в мужские гаметофиты, другие – в женские. Спорофитное поколение начинается с сингамии, дающей диплоидную зиготу. Подвижным сперматозоидам для достижения яйцеклетки требуется водная среда. Внутри калиптры, или брюшка, архегония зигота развивается в зародыш, или молодой спорофит. По мере роста зародыша калиптра тоже увеличивается в размере. В конечном итоге созревающий спорофит разрывает калиптру, вынося спорангий во внешнюю среду. Спорофит в течение всей жизни прикреплен к гаметофиту расширенной стопой.
39. ЛИСТОСТЕБЕЛЬНЫЕ МХИ (настоящие мхи), самый крупный класс моховидных. Включает ок. 14,5 тыс. видов. Широко распространены в холодных и умеренных зонах обоих полушарий – от полярных пустынь Арктики до участков среди антарктических льдов. Многие виды отличаются высокой жизнеспособностью (выдерживают длительное высыхание, а воздушно-сухие споры переносят в течение нескольких часов температуру –200 °C). Листостебельные мхи всегда расчленены на стебель и листья. Листья расположены спирально и собраны на верхушке в виде почки. Различают мхи ортотропные – с вертикальным ростом стебля и плагиотропные – с горизонтальным. В зависимости от роста стебля расположены гаметангии (верхоплодные и бокоплодные мхи). На любой части стебля могут возникнуть ризоиды – волосковидные образования, служащие для прикрепления к субстрату и адсорбции воды. У ряда мхов лист состоит из двух типов клеток: зелёных, осуществляющихфотосинтез, и мёртвых, бесцветных водоносных клеток. Размножение в основном вегетативное. При разрастании моховой дерновины нижняя часть разветвлённого материнского растения отмирает, а верхние части представляют собой обособленные особи. Листостебельные мхи часто играют ведущую роль в растительных сообществах (моховые тундры, замоховелые леса). Особенно велика их роль в формировании болот, где создаются огромные торфяные залежи. Встречаются листостебельные мхи в степях, на скалах, в водоёмах.
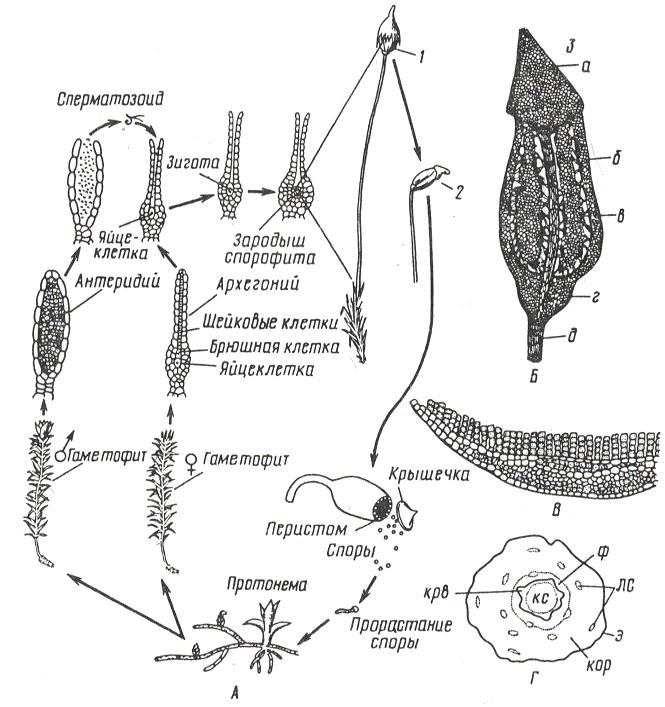
Рис. 11. 2. Кукушкин лен: А – цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы). Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии. Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.