

Рост и развитие растений. Гормональная система растений
Рост и развитие — неотъемлемые свойства любого живого организма. Это интегральные процессы. Растительный организм поглощает воду и питательные вещества, аккумулирует энергию, в нем происходят многочисленные реакции обмена веществ, в результате чего он растет и развивается. Процессы роста и развития тесно взаимосвязаны, так как обычно организм и растет, и развивается. Однако темпы роста и развития могут быть разными, быстрый рост может сопровождаться медленным развитием или быстрое развитие медленным ростом. Так, например, растение хризантемы в начале лета (длинный день) быстро растет, но не зацветает, следовательно, развивается медленно. Подобное происходит с высеянными весной озимыми растениями: они быстро растут, но не переходят к репродукции. Из этих примеров видно, что критерии, определяющие темпы роста и развития, различны. Критерием темпов развития служит переход растений к воспроизведению, к репродукции. Для цветковых растений это закладка цветочных почек, цветение. Критерии темпов роста обычно определяют скоростью нарастания массы, объема, размеров растения. Сказанное подчеркивает нетождественность этих понятий и позволяет рассмотреть процессы роста и развития последовательно.
Существует много определений роста, однако ни одно из них полностью не может считаться удовлетворительным. Прежде всего рост — это необратимое увеличение размеров, объема, массы организма. Однако Д.А. Сабинин справедливо указывал, что это определение неполно и неточно. Дело в том, что у растений увеличение в процессе роста уже имеющихся органов все время сопровождается образованием новых органов (листьев, побегов). Иначе говоря, рост растений включает формообразовательные процессы. При этом вновь появляющиеся побеги, листья качественно отличаются друг от друга. Таким образом, рост растений нельзя рассматривать как чисто количественный процесс. Сказанное позволяет дать следующее определение этого процесса. Рост — это необратимое увеличение объема, массы растений, сопровождаемое новообразованием элементов структуры организма. Под элементами структуры понимают органы, ткани, клетки, а также отдельные клеточные органеллы. В отличие от животных организмов растения в течение всей жизни растут и образуют новые клетки, хотя обычно с некоторыми перерывами (период покоя). Из приведенного определения роста видно, что дать общий критерий, который бы позволил определить темпы этого процесса, очень трудно. В качестве критериев можно брать:
1) высоту и толщину (для стебля);
2) площадь (для листьев и стеблей);
3) массу (сырую и сухую);
4) число клеток;
5) содержание белка;
6) содержание ДНК и др.
Однако ни один из этих показателей не дает полной картины процесса роста. Нередко в зависимости от выбранного критерия мы получаем различные, а иногда и прямо противоположные результаты. Дело в том, что разные органы растения растут с различной скоростью. Не всегда увеличение высоты растения сопровождается увеличением массы, и изменение сырой массы не всегда аналогично изменению сухой. Нередки случаи, когда параметры роста изменяются прямо противоположным образом. Так, например, при прорастании семян в темноте увеличивается объем, но сухая масса уменьшается. Этиолированные проростки интенсивно растут в длину, но масса их не увеличивается. В покоящихся растениях идет скрытый рост, который выражается в новообразовании элементов структуры и может не сопровождаться увеличением массы и даже объема. В силу сказанного, говоря о росте, необходимо уточнять, что именно имеется в виду — увеличение длины или массы, числа клеток или их размеров.
Типы роста органов растения
Характерной чертой ростовых процессов растительных организмов является их локализация в определенных тканях — меристемах. Меристемы различны по расположению в отдельных органах. Апикальные, или верхушечные, меристемы расположены в окончаниях (верхушках) стебля и корня. Вставочные, или интеркалярные, меристемы характерны для стебля (рост междоузлий у злаков) и для некоторых листьев. Латеральные меристемы обеспечивают рост стебля в толщину (камбий, феллоген).
Клетки меристемы делятся; дочерние клетки достигают размеров материнской и снова делятся. Однако размер и объем меристем остаются постоянными. Это связано с тем, что большинство меристематических клеток через несколько делений (4—5) переходят к росту растяжением. Однако есть инициальные клетки, которые делятся в течение всей жизни органа. Показано также, что апикальные меристемы корня и стебля состоят из двух типов клеток, резко отличающихся по функциям и по способности к делению. Меристематические клетки, расположенные на самом верху стебля или корня, не прекращают делиться в течение всего периода роста. Эту зону меристематических клеток называют покоящимся центром (для корня) или меристемой ожидания (для стебля). Более длительная способность к делению является следствием меньшей частоты делений и большей длительности интерфазы. Одновременно данные клетки характеризуются и большей длительностью митотического цикла. Вместе с тем эти клетки более устойчивы к неблагоприятным воздействиям. Так, в них реже возникают хромосомные аберрации, что очень важно для сохранения жизнеспособности организма (В.Б. Иванов). Клетки меристемы ожидания стебля менее дифференцированы, что облегчает их последующую дифференциацию и имеет значение при переходе к образованию генеративных органов. Уже на первой фазе роста — фазе деления — клетки, находящиеся в нижней части меристемы, начинают дифференцироваться. В них постепенно накапливаются физиологические, а затем и морфологические различия. Эти различия обусловлены местоположением клетки, взаимодействием ее с другими клетками, а также той генетической программой, которая в нее вложена.
Вопрос о причинах дифференциации клеток — один из наиболее сложных вопросов физиологии. Все клетки данного организма обладают одинаковым геномом, а следовательно, все клетки должны обладать и одинаковыми потенциальными возможностями (тотипотентностью). Тотипотентность проявляется в способности дифференцированных растительных клеток превращаться в эмбриональные (дедифференцированные) и при определенных условиях давать начало целому организму. В частности, тотипотентность клеток проявляется в широко распространенной способности растительного организма к восстановлению утраченных частей — регенерации. Хорошо известным примером регенерации является образование придаточных корней при черенковании растений. Черенками называется любая часть растения, стебля, корня, листа, которая, будучи отделена; способна дать новую особь. После отделения черенка клетки, прилегающие к поверхности среза, начинают усиленно делиться и образуется наплыв, или каллюс. Вначале каллюс — это недифференцированная ткань, а затем в ней обособляются проводящие элементы и закладываются корневые зачатки, из которых развиваются придаточные корни. Однако наиболее ярко тотипотентность клеток проявляется при культуре изолированных тканей.
Развитие растений
На протяжении всей жизни, т. е. онтогенеза, растение растет и развивается. Развитие — ход качественных последовательных изменений структуры, который проходит организм от возникновения из оплодотворенной яйцеклетки до естественной смерти. Общая схема развития каждого организма запрограммирована в его наследственной основе. Растения резко различаются по продолжительности жизни. Известны растения, которые заканчивают свой онтогенез на протяжении 10—14 суток (эфемеры). Вместе с тем существуют растения, продолжительность жизни которых исчисляется тысячелетиями (секвойи). Независимо от продолжительности жизни все растения можно разделить на две группы: монокарпические, или плодоносящие один раз, и поликарпические, или плодоносящие многократно. К монокарпическим относят все однолетние растения, большинство двулетних, а также некоторые многолетние. Многолетние монокарпические растения (например, бамбук, агава) приступают к плодоношению после нескольких лет жизни и после однократного плодоношения отмирают. Большинство многолетних растений относят к поликарпическим.
Этапы развития растении
Развитие каждого растительного организма, так же как и животного, расчленимо, т. е. оно проходит ряд этапов. Эти этапы характеризуются морфологическими и физиологическими признаками. Физиологические критерии этапов развития изучены слабо. При выделении этапов используют, главным образом, морфологические признаки. Для семенных растений можно выделить следующие этапы развития (М.Х. Чайлахян):
1) эмбриональный — от оплодотворения яйцеклетки до прорастания зародыша. Этот этап можно разделить на два периода:
а) эмбриогенез — период, в котором эмбрионы находятся на материнском растении;
б) покой — период от конца формирования семени и до его прорастания;
2) молодости (ювенильный) — от прорастания зародыша до закладки цветочных зачатков, характеризующийся усиленным ростом;
3) зрелости — от момента закладки цветочных зачатков до оплодотворения (появления новых зародышей);
4) размножения— от оплодотворения до полного созревания семян;
5) старости — от периода созревания семян до отмирания.
Поликарпические растения этапы эмбриональный и молодости проходят один раз, однако часто эти этапы характеризуются большой продолжительностью (несколько лет). Этапы зрелости и размножения наступают один раз, но осуществляются многократно. Этап старости у поликарпических растений также может продолжаться несколько лет. На протяжении каждого из перечисленных этапов развития в растении возникают новые органы. Процесс формирования этих органов называют органогенезом. Этот процесс можно изучить путем микроскопических наблюдений за дифференциацией верхушечных меристем и изменениями в формирующихся органах. Для одно- и двулетних растений выделено 12 последовательных этапов органогенеза. При этом на I и II этапах происходит дифференциация вегетативных органов, на III и IV—дифференциация зачаточного соцветия, на V—VIII — формирование цветков, на IХ — оплодотворение и образование зиготы, на X— XII — рост и формирование семян (Ф.М. Куперман). Основой морфологических, структурных изменений является изменение физиолого-биохимических процессов. Именно поэтому, как правило, внутренние физиолого-биохимические изменения предшествуют морфологическим. Однако проявляется и обратная зависимость. Вновь появившиеся структуры, органы оказывают влияние на уровень и направленность процессов метаболизма. Таким образом, можно считать, что есть единый процесс физиолого-биохимических и морфологических изменений. При этом физиолого-биохимические изменения, определяющие наступление последующего этапа, происходят в структурах, образовавшихся на предыдущем этапе развития организма. Старение и смерть как этапы программы онтогенеза. Старение — активный процесс развития, который зашифрован в генетической программе и регулируется специфическими сигналами или импульсами, возникающими под влиянием условий среды. Во время процессов старения экспрессия большинства генов затухает.
Однако экспрессия некоторых генов наоборот возрастает. Активируются гены, которые кодируют белки-ферменты, вызывающие процессы распада (протеазы, нуклеазы, липазы, ферменты, разрушающие хлорофилл и др.). Программированная смерть у растений в отличие от животных изучена мало. Показано, что программированная смерть клетки вызывается специальными сигналами и в свою очередь вызывает экспрессию ряда генов. Программированная смерть, сопровождаемая определенными морфологическими и биохимическими изменениями, суммируется как апоптоз — генетически детерминированная смерть клеток, которая является обязательной частью развития клеток. Показано, что апоптоз у растений сходен с таковым у животных, но имеются структурно-морфологические различия, связанные с наличием у растений вакуоли и клеточной стенки. Апоптоз может проявляться на самых различных органах и тканях организма: колеоптили, лепестки цветка, корни, эндосперм и другие. При апоптозе происходит ряд изменений: реорганизуется цитоплазма, изменяется фрагментация ДНК, наблюдается распад ядра. Апоптоз зависит от ряда внешних и внутренних факторов (инфекции, различные стрессоры). Важнейшая функция запрограммированной смерти клеток для растений — защита от патогенных организмов. При заражении в клетках быстро накапливаются фенольные соединения, и они отмирают. Это проявляется в образовании на органах растения круглых пятен мертвых клеток — некрозов. Роль некротических пятен заключается в изоляции токсических веществ для защиты здоровых органов. Возникновение некрозов является формой запрограммированной смерти клеток. На мутантах арабидопсиса показано, что под влиянием инфекции в них происходит каскад растворений и появляются некротические пятна. Непосредственной причиной смерти клеток у мутантов является аккумуляция перекисных соединений кислорода. Апоптоз регулируется гормональной системой, что связано с контролем над метилированием ДНК. Так, показано, что АБК стимулирует изменения ДНК, происходящие при апоптозе. Существуют данные, что процесс деметилирования ДНК ответственен за индукцию генов апоптогенных белков или за репрессию генов антиапоптозных генов (Б.Ф. Ванюшин). Наряду с апоптозом существует генетически запрограммированная смерть органов и организма в целом. Так, листья генетически запрограммированы для старения и смерти. Предложено называть процессы гибели отдельных органов органоптозом, а целого организма феноптозом (В.П. Скулачев). Запрограммированную смерть можно наблюдать при дифференцировке трахеид, когда разрушаются ядро и хроматин. После единственного акта цветения заканчивается жизнь у всех монокарпических растений.
Гормоны роста растений (фитогормоны)
Для многоклеточных организмов характерен тип регуляции, который связан с взаимодействием между отдельными клетками, тканями или даже органами. Для осуществления такой координации в организме вырабатываются гормоны. Гормоны растений получили название фитогормонов. Фитогормоны — это вещества, вырабатывающиеся в процессе естественного обмена веществ и оказывающие в ничтожных количествах регуляторное влияние, координирующее физиологические процессы. В этой связи к ним часто применяется термин — природные регуляторы роста. В большинстве случаев, но не всегда фитогормоны образуются в одних клетках и органах, а оказывают влияние на другие. Иначе говоря, гормоны способны к передвижению по растению и их влияние носит дистанционный характер. Большинство физиологических процессов, в первую очередь рост, формообразование и развитие растений, регулируется гормонами. Гормоны играют ведущую роль в адаптации растений к условиям среды. Известны следующие пять групп фитогормонов: ауксины, гиббереллины, цитокинины, абсцизовая кислота, газ этилен. В последнее время к ним относят брассины (брассиностероиды). Условно можно отнести первые три группы—ауксины, гиббереллины и цитокинины и частично брассины — к веществам стимулирующего характера, тогда как абсцизовую кислоту и этилен — к ингибиторам.
Ауксины — соединения преимущественно индольной природы: индолилуксусная кислота и ее производные (индолил-3-ацетальдегид, индолил-3-ацетонитрил, индолил-3-молочная кислота, метиловый и этиловый эфиры ИУК, триптамин и др.). Ауксин образуется из триптофана, который синтезируется из шикимовой кислоты, возникающей в процессе дыхания из ФЭП (из гликолиза) и Эр-4-Ф (из пентозофосфатного цикла) по схеме: ФЭП → Эр-4-Ф → шикимовая кислота → триптофан → ИУК (рис. 115). Превращение триптофана в ИУК усиливается под действием гиббереллина и уменьшается под влиянием хлорхолинхлорида, этилена и радиоактивного облучения.
Гиббереллины являются тетрациклическими карбоновыми кислотами — гибберелловая кислота (ГА) и другие гиббереллины. Известно более 70 гиббереллинов, различающихся деталями строения. Наиболее распространенным и изученным является гиббереллин А3 или гибберелловая кислота (ГА3). Гиббереллины образуются из мевалоновой кислоты, синтезированной из ацетил~КоА: ацетил ~КоА → мевалоновая кислота → гиббереллин (рис. 116).
Цитокинины — производные 6-аминопурина, синтезируются посредством конденсации аденозин-5-монофосфата и изопентенилпирофосфата. Предшественником цитокининов являются мевалонат и пурин. Основным природным цитокинином служит зеатин, выделенный из семян кукурузы молочной спелости.
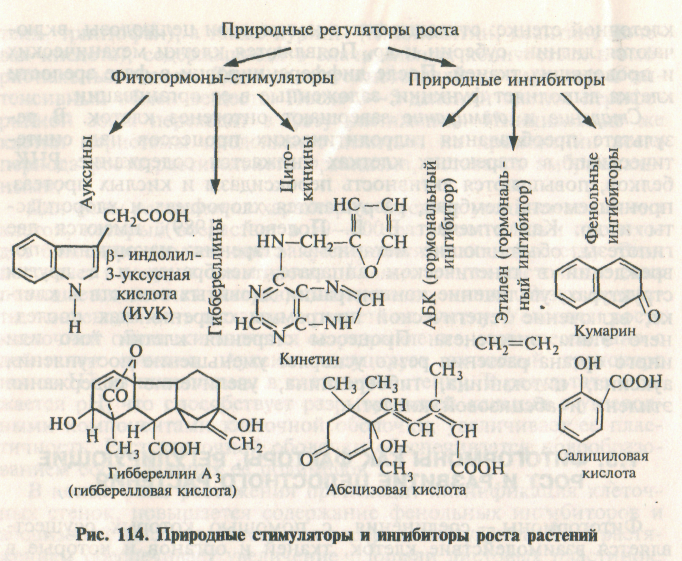
Абсцизовая кислота (АБК) и ее производные — соединения терпеноидной природы. АБК синтезируется двумя способами: из мевалоновой кислоты через изопентилпирофосфат и геранилпирофосфат; в результате деградации каротиноидов, в частности виолоксантина, через ксантоксин. В первом случае АБК образуется из тех же предшественников (ацетат и мевалонат), что и гиббереллины. Биосинтез АБК ингибируют цитокинины и ингибиторы биосинтеза гиббереллинов.
Этилен — гормональный фактор, газ Н2С=СН2. Этилен синтезируется из метионина. Ближайший предшественник этилена — 1-аминоциклопропан-1-карбоновая кислота (АЦК).
Фенольные ингибиторы — вещества фенольной природы: кофейная, хлорогеновая, коричная кислоты и др. Эти вещества не являются фитогормонами. Они не передвигаются по сосудам и действуют в концентрациях, в 2—3 раза больших, чем гормоны. Общим для всех предшественников фенольных соединений является наличие шикимовой кислоты. Дальнейшие ее превращения приводят к образованию фенилаланина и тирозина, а затем коричной и паракумариновой кислот, дающих начало всему разнообразию фенольных веществ (около 2000).
Таким образом, следует подчеркнуть: все регуляторные соединения синтезируются из аминокислот или органических кислот; их биосинтез происходит из продуктов фотосинтеза и дыхания; общие предшественники (хорезмовая, мевалоновая кислоты), от которых возникают биосинтетические пути, направленные на образование как стимуляторов, так и ингибиторов, являются ключевыми.
Фитогормоны полифункциональны, регулируют многие физиологические процессы, физиологическое действие их на растение зависит от следующих факторов:
специфики фитогормона — спектра физиологических действий на растительный организм данного фитогормона;
специфики объекта — видовых, органных, тканевых, возрастных и других особенностей растительного объекта, определяющих его восприимчивость к фитогормону;
концентрации фитогормона — определенных границ концентраций, в которых фитогормон активирует или ингибирует данный физиологический процесс или функцию;
соотношения данного фитогормона с другими — от функционирования многокомпонентной гормональной системы, составляющей гормональное поле растения;
обеспеченности растительного объекта необходимыми факторами минерального и углеродного питания;
эндогенного содержания фитогормона — ответной реакции растительного объекта на экзогенный гормон, зависящей от его концентрации внутри растения;
напряженности факторов внешней среды (свет, температура, вода и др.), создающих необходимые условия для действия фитогормона.
Физиологические функции фитогормонов. Действуя на растение экзогенными гормонами, наблюдают возникшие эффекты и судят о роли фитогормонов.