1.2.Биосинтез жирных кислот и построенных из них фосфолипидов
Существует два основных пути биосинтеза липидов. Первый путь основан на синтезе жирных кислот из ацетилкофермента А с дальнейшим превращением их в жиры, воска, фосфолипиды и некоторые другие более специализированные биологически активные вещества, например в простагландины. Второй путь имеет в своей основе синтез изопентенильных производных, из которых образуются многочисленные соединения с разветвленной цепью и циклические структуры, в том числе различные терпены и стероиды. Этот путь рассматривается в следующем параграфе.
Биосинтез жирных кислот в главных своих чертах представляет собой обращение пути окислительной деструкции жирных кислот. Он приводит к кислотам, содержащим четное число углеродных атомов. У эукариот ацетилкофермент А преимущественно образуется в митохондриальном матриксе, в то время как биосинтез жирных кислот проходит в цитоплазме. Поэтому необходим транспорт активированных ацетильных остатков через митохондриальные мембраны. Для самого ацетилкофермента А эти мембраны непроницаемы. Поэтому транспорт проходит с помощью вспомогательного переносчика, роль которого играет карнитин. Как в матриксе, так и в цитоплазме осуществляется перенос ацетильного остатка между коферментом А и карнитином, катализируемый специальным ферментом карнитин ацетилтрансферазой.
В отличие от окислительной деструкции, которая происходит с ацильными остатками, связанными с коферментом А, сборка ацильных остатков происходит на специальном небольшом белке, носящем название ацилпереносящий белок, который в дальнейшем обозначается как АСР (acyl carrier protein) или АСР–SH. Его функциональной группой, как и в случае кофермента А, является SH-группа фосфопантотеина, связанного с белком фосфоэфирной связью через гидроксигруппу серина.

После образования в цитоплазме ацетилкофермента А часть ацетильных остатков переносится на АСР по реакции
![]()
катализируемой АСР-ацетилтрансферазой.
Параллельно с этим происходит карбоксилирование ацетилкофермента А по реакции

приводящие к образованию малонилкофермента А. Образовавшийся малонильный остаток также переносится на ацил-переносящий белок:
![]()
Удлинение цепи начинается с переноса ацетильного фрагмента на малонил-АСР:
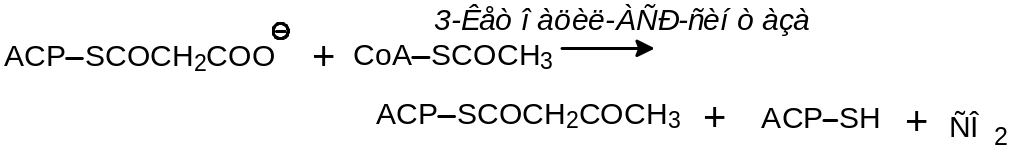
Последующие стадии удлинения углеродной цепи представляют собой обращение соответствующих реакций окислительной деградации
![]()
![]()
Цикл завершается процессом:
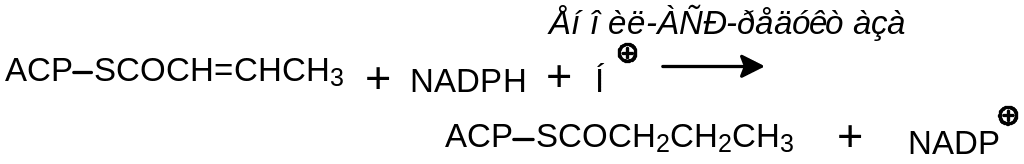
По завершении этого первого цикла осуществляется перенос бутироильного остатка на малонил-АСР:
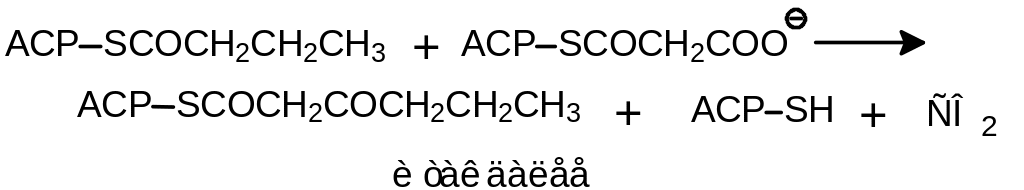
и начинается новая серия реакций, приводящая в итоге к восстановлению следующей кетогруппы.
Общий баланс одного цикла можно записать в виде:
![]()
Вторым важным компонентов жиров и важных фосфолипидов является глицерин. Источником глицеринового фрагмента является дигидроксиацетонфосфат, который восстанавливается с помощью NADH в реакции, катализируемой глицерин-3-фосфат дегидрогеназой. Далее проходит двухступенчатое Ацилирование обеих гидроксигрупп с использованием ацилкофермента А:
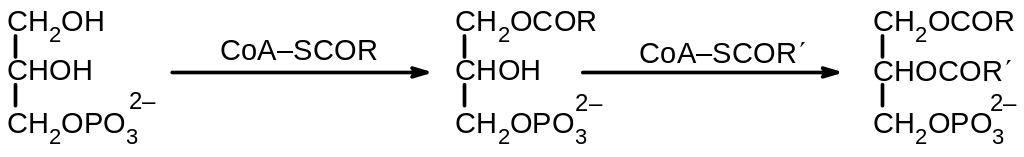
Итогом этих реакций являются фофтатиды. Далее превращения могут идти двумя основными путями.
Один путь начинается с гидролиза фосфатида и приводит к образованию 1,2-диацилглицеринов. Последние могут ацилироваться ацилкоферментом А с образованием триацилглицеридов, т.е. жиров.
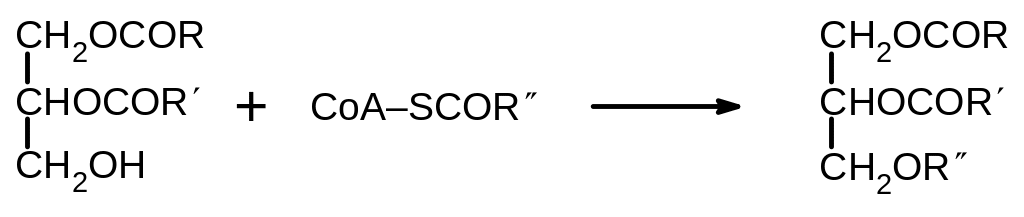
На этом же пути могут образовываться важнейшие Фосфолипиды — фосфатидилэтаноламин и фосфатидилхолин. Эти процессы происходят с участием производных цитидинфосфата:
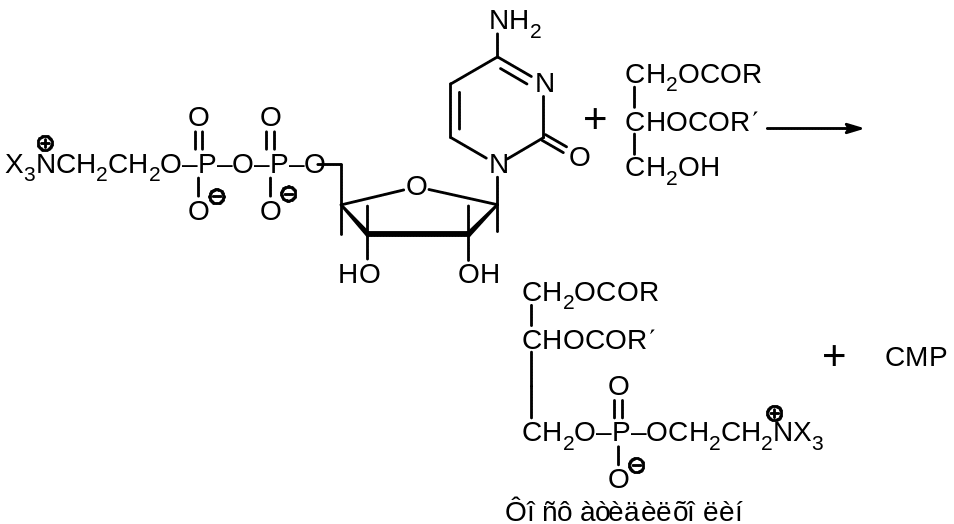
Второй путь биосинтеза фосфолипидов исходит из фосфатида, который взаимодействует с СТР с образованием СDР-диглицерида:
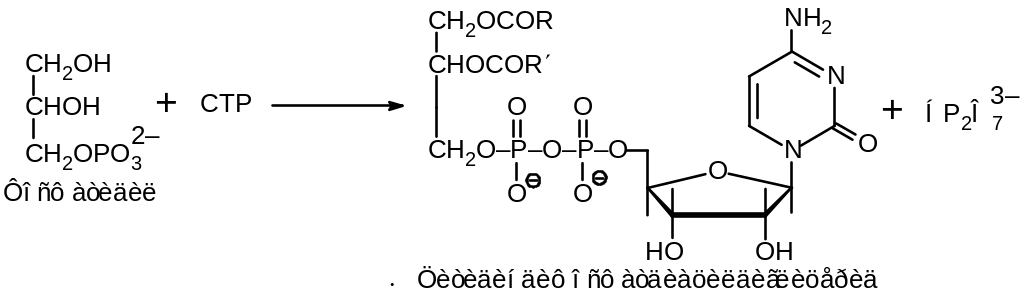
Образовавшееся производное CDP реагирует со спиртовыми группами, например:

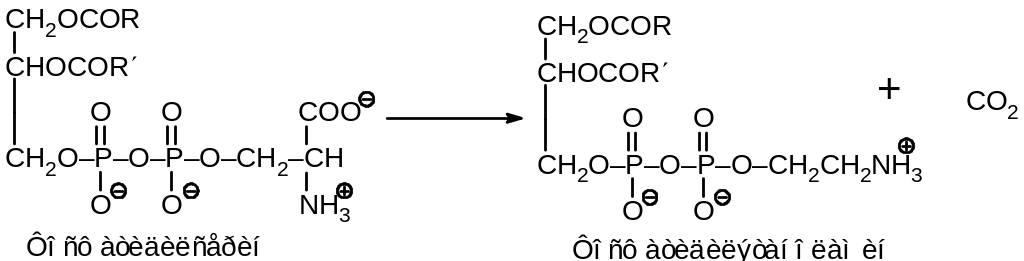
При взаимодействии с серином получается фосфатидилсерин. Последний может подвергаться декарбоксилированию с образованием фосфатидилэтаноламина: