

Устойчивое равновесие популяции (популяционный гомеостазис)
Смертность и рождаемость величины противоположные. При нарастании плотности населения смертность обычно увеличивается, рождаемость падает. Очевидно, что должна существовать и плотность, при которой кривые рождаемости и смертности пересекаются (рис. на доске).
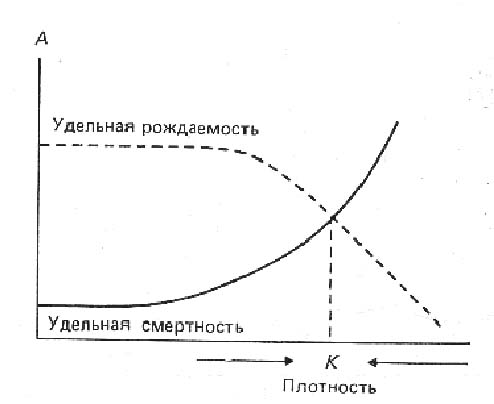
Такая плотность (точка К на рисунке) отвечает состоянию устойчивого равновесия популяции: к данному значению будут стремиться все значения плотности. Слишком низкая плотность популяции будет способствовать снижению смертности и увеличению рождаемости, слишком высокая плотность – увеличению смертности и снижению рождаемости. Таким образом, внутривидовая конкуренция, влияя на рождаемость и смертность, регулирует плотность населения и численность популяции, поддерживая ее на стабильном уровне, при котором рождаемость уравновешена смертностью. Такую плотность называют предельной плотностью насыщения. Поскольку эта величина характеризует тот размер популяции, который могут поддержать имеющиеся ресурсы среды, предельная плотность насыщения может рассматриваться как показатель емкости среды (carrying capacity, capacité d’acceuil), т.е. того количества организмов, которое способно жить в данных условиях без особого угнетения. Зависимость смертности, выживаемости и рождаемости от плотности популяции нередко называется аторегуляцией, саморегуляций, популяционной регуляцией и т.д.
Принцип конкурентного исключения (или принцип Гаузе)
Суть принципа конкурентного исключения заключается в том, что каждый вид имеет свою собственную экологическую нишу, при этом разные виды не могут занимать одну и ту же нишу. Независимо от форм межвидового взаимодействия, результатом использования разными видами одной и той же ниши будет вытеснение одним видом другого.
Правило это было сформулировано Г.Ф.Гаузе (1934) на основании эмпирических наблюдений за двумя видами инфузорий: Paramecium caudatum и P. aurelia. Его эксперименты показали, что в одновидовых культурах нарастание численности популяций обоих видов шло по классической S-образной кривой. Однако в смешанных культурах по такому типу развивалась популяция лишь одного из видов, другая популяция вымирала. Обобщенный результат этого эксперимента гласит, что виды, конкурирующие за общий дефицитный ресурс, не могут входить в состав одного сообщества (или биоценоза). На доске пояснить S-образную кривую динамики численности.
Со времен работы Г.Ф.Гаузе в это правило внесено много дополнений и исправлений. В частности уточнялось, что «полные конкуренты не могут сосуществовать бесконечно». Как бы то ни было, это правило до сих пор является одним из главных догматов теоретической экологии: если два вида сосуществуют (используя один и тот же дефицитный ресурс), то между ними должно быть какое-то экологическое различие, а это означает, что каждый из них занимает свою особую нишу.
Если же конкуренция все же зарегистрирована, то считается, что, конкурируя с более сильным видом, слабый конкурент утрачивает свою реализованную нишу. То есть, выход из конкуренции реализуется через изменения собственных требований к среде одним из конкурентов. Меняется его «образ жизни», он занимает пусть и близкую, но несколько отличную новую экологическую нишу. Если же слабейший конкурент оказывается неспособным к таким изменениям собственной ниши, его будущее выглядит печальным: в соответствие с принципом конкурентного исключения он должен вымереть. Таким образом, речь идет о разграничении экологических ниш разных видов, которые в прошлом могли использовать одну нишу. Лишь в этом случае разные виды приобретают способность сосуществовать в одном биоценозе (сообществе).
Современный подход к проблеме разделения экологической ниши несколькими видами не столь категоричен. Он допускает, что в некоторых случаях два разных вида могут сосуществовать, разделяя при этом одну и туже экологическую нишу и находясь при этом в состоянии межвидовой конкуренции. Например, Э.Хейрстон, изучая два вида сухопутных саламандр Plethodon glutinosus и Plethodon jordani в южной части Аппалачских гор в США, отметил, что в некоторых районах зоны их обитания перекрывались. Отмечено также, что исходно оба вида испытывали неблагоприятное воздействие со стороны друг друга. После удаления одного из видов, у оставшегося наблюдалось увеличение численности и/или плодовитости и/или выживаемости. Следовательно, в местах совместного обитания эти виды хотя и конкурировали друг с другом, но все же сосуществовали.
Очевидно, что правило конкурентного исключения вряд ли можно воспринимать как абсолютное. Если его принять в той формулировке, что виды, занимающие общую экологическую нишу взаимоисключают друг друга, то встает вопрос о том, что принимается под самой нишей.
- Если нишу обозначить как «экологическое пространство вида, в котором он не имеет конкурентов» правило Г.Ф.Гаузе становится тавтологическим и теряет смысл.
- Если речь идет просто о конкуренции за ресурс, реальное положение таково, что становление биоценотических систем связано с расхождением ниш и снижением уровня конкуренции. В таком варианте правило конкурентного исключения подразумевает пространственное (а иногда и функциональное) разобщение видов в составе биоценоза.
Абсолютное вытеснение, изучая современный состав экосистемы, зафиксировать практически невозможно. Пожалуй, только изучение островных фаун дает примеры этого процесса. Так, в комплексах островов конкурирующие виды не встречаются вместе, хотя и заселяют близко расположенные острова. Например, на островах Средиземного моря встречаются ящерицы лишь одного из двух близких видов, но никогда не представители обоих видов одновременно.
Кроме того, в реальности равновесие между конкурирующими видами может неоднократно нарушаться, и преимущество будет переходить от одного вида к другому; следовательно, сосуществование возможно просто благодаря изменению условий среды. И в этом случае принцип Гаузе просто не применим. Популярным примером этого является «Планктонный парадокс» (впервые его привел Дж.Хатчинсон в 1961 г.).
Парадокс этот состоит в том, что многочисленные виды планктонных организмов часто сосуществуют в простой среде, где, по-видимому, мало возможностей для разделения ниш. Эта среда, хотя она и очень проста, постоянно претерпевает различные изменения, в частности суточные или сезонные. В любой отдельный промежуток времени условия среды могут способствовать вытеснению определенного вида, однако эти условия меняются, и еще до того, как данный вид окажется окончательно вытесненным, они могут сложиться вполне благоприятно для его существования. Другими словами, исход конкурентных взаимодействий в равновесном состоянии может НЕ играть решающей роли, если условия среды изменяются еще до того, как достигается равновесие. А поскольку любая среда изменчива, равновесие между конкурентами должно постоянно сдвигаться и сосуществование нередко будет наблюдаться при таком разделении ниш, при котором в стабильных условиях происходило бы исключение одного из видов.
Завершая обсуждение конкуренции, следует непременно упомянуть еще о двух закономерностях:
Биоценотическое правило В.С.Ивлева (1955), согласно которому межвидовое напряжение гораздо значительнее, чем внутривидовые отношения. Поэтому внутривидовая конкуренция более вероятна (и ярче выражена), чем межвидовая.
В многовидовых сообществах не исключено возникновение «диффузной» конкуренции, т.е. «войны всех против всех» (MacArthur, цит. по Шенброт 1986). Ее возникновение возможно в результате некой «переадресации» конкуренции с одного вида на другой. Теория диффузной конкуренции не разработана.