

Зрачковый рефлекс. Проекция зрительного поля на сетчатку. Бинокулярное зрение. Конвергенция глаз. Дивергенция глаз. Поперечная диспарация. Ретинотопия. Регуляция интенсивности светового потока
Приспособление к изменяющимся условиям освещения происходит с помощью зрачкового рефлекса. Ширина зрачка уменьшается при ярком свете благодаря сокращению кольцевых мышц, управляемых парасимпатическими волокнами глазодвигательного нерва, а при слабом освещении зрачок расширяется с помощью радиальных мышц, получающих симпатическую иннервацию. Уменьшая просвет зрачка, глаз защищается от избытка света, а увеличивая ширину зрачка, он повышает чувствительность зрительной системы к воспринимаемым стимулам. Сужение зрачков повышает глубину резкости, что позволяет лучше видеть удаленные предметы. Прирасширении зрачков глубина резкости снижается, а вместе с ней снижается острота зрения, которая характеризуется максимальной способностью глаза различать две соседние точки зрительного пространства как отдельные. В норме глаз различает две точки, видимые под углом в одну минуту при достаточно ярком освещении.
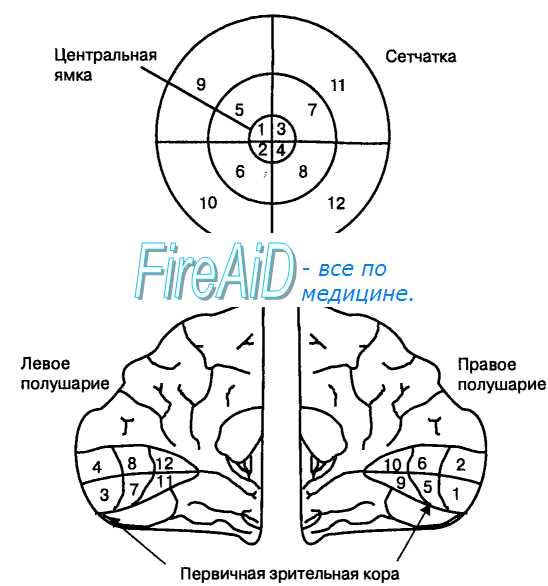
Рис. 17.6. Ретинотопическая проекция сетчатки в первичную зрительную кору. Пространственное соотношение между объектами зрительного поля соблюдается в его проекции на соответствующие друг другу (корреспондирующие) области сетчатки каждого глаза. Афферентные сигналы от носовых половин сетчатки благодаря частичному перекресту волокон зрительных нервов поступают в противоположное полушарие. Афферентные сигналы от височных половин сетчаток попадают с помощью неперекрещивающихся волокон зрительных нервов в ипсилатеральное полушарие. В связи с этим в каждое полушарие поступает зрительная информация о противоположной ему половине зрительного поля и с сохранением его пространственных координат (принцип ретинотопии). При направленном на определенные объекты внимании световые лучи, отраженные от них, всегда фокусируются в центральной ямке. Поэтому центральная ямка и прилегающие к ней области сетчатки имеют наибольшую площадь представительства в первичной зрительной коре.
Проекция зрительного поля на сетчатку
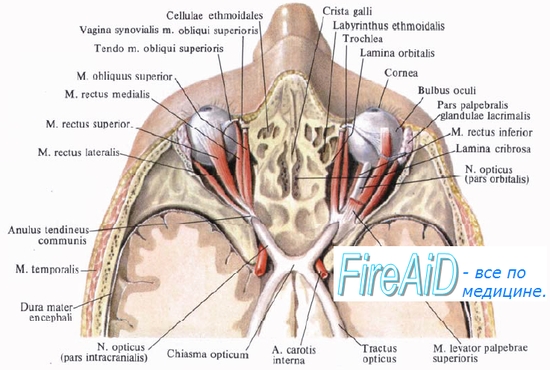
Благодаря механизму аккомодации глаз представляет собой самофокусирующуюся оптическую систему, позволяющую получать на сетчатке четкое изображение приближенных и удаленных предметов. Кроме того, глаз подвижен, что позволяет ему следить за перемещающимися объектами. Движения глаза обеспечиваются сокращениями глазодвигательных мышц (двух косых и четырех прямых), которые позволяют повернуть глаз внутрь или наружу, вверх или вниз. Оба глаза поворачиваются бинокулярно, т. е. содружественно, и при рассмотрении близких предметов глазные оси сходятся (конвергенция глаз), а при рассмотрении далеких предметов — расходятся (дивергенция). Благодаря бинокулярному зрению каждая точка зрительного поля проецируется на соответствующие друг другу участки сетчатки обоих глаз. Изображение одного и того же предмета на сетчатках левого и правого глаза немного различается, поскольку глаза занимают разную позицию по отношению к зрительному полю. Разница изображений наблюдаемого предмета на сетчатке левого и правого глаза обозначается термином поперечная диспарация, она способствует последующему преобразованию двухмерного изображения на сетчатке в трехмерное изображение, создаваемое в процессе переработки зрительной информации в коре.
Зрительное поле представляет собой область, видимую обоими глазами в тот момент, когда они и голова неподвижны. При таком условии левая половина зрительного поля проецируется на носовую половину сетчатки левого глаза и височную половину сетчатки правого глаза. Соответственно, правая половина зрительного поля проецируется на носовую половину сетчатки правого глаза и височную половину сетчатки левого глаза. Поскольку оптическая система глаза перевертывает изображение, верхняя половиназрительного поля проецируется на нижнюю половину сетчатки, а нижняя половина зрительного поля — на верхнюю половину сетчатки. При дальнейшей переработке информации в наружном коленчатом теле и первичной зрительной коре сохраняются все пространственные координаты зрительного поля: каждая область сетчатки связана с соответствующим представительством в коре, что называется ретинотопией (рис. 17.6).
Движения глаз. Следящие движения глаз. Быстрые движения глаз. Центральная ямка. Саккадамы.
При наблюдении за движущимися в зрительном поле объектами, а также при движении человека относительно окружающего мира происходят следящие движения глаз, благодаря которым изображение в одной и той же области сетчатки сохраняется неизменным. При зрительном восприятии неподвижных объектов, имеющих многочисленные детали формы, а также во время чтения происходятбыстрые движения глаз, предназначенные для фиксации наиболее информативных деталей объекта. Например, при восприятии человеческого лица фиксируются его контуры, но с наибольшим вниманием рассматриваются глаза и рот, изображения которых глаз проецирует не вообще на сетчатку, а на ту ее область, которая обладает максимальной разрешающей способностью. Это центральная ямка, или fovea, представляющая собой небольшое углубление диаметром около 3 мм в центре сетчатки.
При рассматривании любых объектов глаза ежесекундно совершают около трех очень быстрых непроизвольных и субъективно не ощущаемых движений, которые называются саккадами. Благодаря таким движениям изображение на сетчатке регулярно смещается, вызывая раздражение разных фоторецепторов. Необходимость саккад объясняется свойством зрительной системы сильнее реагировать на изменяющееся раздражение (появление или исчезновение стимула), тогда как на постоянную стимуляцию она отвечает слабо. При проецировании на сетчатку постоянного изображения с помощью миниатюрного проектора, прикрепленного к контактной линзе, это изображение перестает восприниматься спустя несколько секунд, и вместо него в зрительном ощущении появляется сплошное серое поле. По истечении нескольких секунд восприятие проецируемого на сетчатку изображения восстанавливается, но затем снова исчезает и т. д.
Преобразование энергии света в сетчатке. Функции ( задачи ) сетчатки. Слепое пятно.
Задняя стенка глаза выстлана эпителием, содержащим черный пигмент меланин, который препятствует отражению света и поглощает его излишки. К пигментному эпителию примыкает слой фоторецепторов, а перед ним расположены слои горизонтальных, биполярных, амакриновых и ганглиозных клеток, чьи аксоны образуют зрительный нерв.
В связи с инверсией сетчатки решаются две задачи: 1) обеспечение максимальной прозрачности клеток, расположенных на пути прохождения света к фоторецепторам, 2) выделение в слое фоторецепторов промежутка для прохождения волокон зрительного нерва.
Решение первой задачи заключается в анатомическом устройстве центральной ямки, где биполярные и ганглиозные клетки сетчатки располагаются вокруг нее, а их относительно прозрачные безмиелиновые волокна не мешают прохождению света к фоторецепторам. В других областях сетчатки световые лучи отчасти рассеиваются при прохождении сквозь тела клеток, расположенных перед фоторецепторами, но в центральной ямке кванты света без помех попадают на фоторецепторы, представленные здесь преимущественно колбочками. При направленном на определенный объект зрительного поля внимании его изображение всегда фокусируется в центральной ямке для обеспечения максимальной остроты зрения.
Сосок зрительного нерва образован аксонами ганглиозных клеток, отсутствие в этой области фоторецепторов делает ее не чувствительной к свету, в связи с чем она получила название слепого пятна. На соски правого и левого глаза благодаря диспарации попадают неодинаковые части проецируемого изображения, поэтому в условиях естественного зрительного восприятия пробел изображения на сетчатке не осознается. Кроме того, быстрые движения глаз позволяют получать нужную информацию о любой области рассматриваемого изображения, и отсутствие сигналов от слепого пятна замещается информацией, поступившей от ближайших к нему фоторецепторов.
Скотопическая система сетчатки ( ночное зрение ). Фотопическая система сетчатки ( дневное зрение ). Колбочки и палочки сетчатки. Родопсин.
Фоторецепторный слой сетчатки человека образован примерно 130 миллионами клеток, из которых около семи миллионов являются колбочками, основная масса которых сосредоточена в области центральной ямки, а все остальные фоторецепторы представлены палочками. У обеих разновидностей фоторецепторов существуют три функциональные области: 1) наружный, или внешний, сегмент, ориентированный в направлении эпителиального пигментного слоя и содержащий зрительный пигмент; 2) внутренний сегмент, в котором расположено клеточное ядро и происходят биохимические процессы, связанные с жизнедеятельностью клетки; 3) синаптические окончания, предназначенные для передачи информации от фоторецепторов к биполярным клеткам с помощью медиатора глутамата.
Зрительный пигмент палочек родопсин состоит из двух компонентов: это молекула ретиналя, образующаяся из витамина А и способная поглощать свет, а также крупная белковая молекула опсина, не поглощающая свет. Молекула опсина представляет собой извитую цепь из 348 аминокислот, которая семь раз проходит через мембрану зрительного диска, образованного из клеточной мембраны фоторецептора. В наружном сегменте фоторецептора имеется большое количество таких дисков, расположенных подобно стопке поставленных друг на друга монет. Ретиналь существует в темноте как 11-цис-ретиналь, такая форма изомера идеально соответствует упорядоченному расположению аминокислот в опсине. Энергия поглощенных фотонов превращает ретиналь в 11-транс-изомер, что приводит к конформационным изменениям молекулы опсина и превращению родопсина в нестабильный метародопсин, который сразу же распадается на ретиналь и опсин. Таким образом, действие света уменьшает концентрацию родопсина в фоторецепторе, что приводит к изменениям активности вторичных посредников и величины мембранного потенциала фоторецептора. В темноте происходит ферментативный ресинтез расщепленного родопсина, для которого используется витамин А, поступающий в организм человека с пищей.
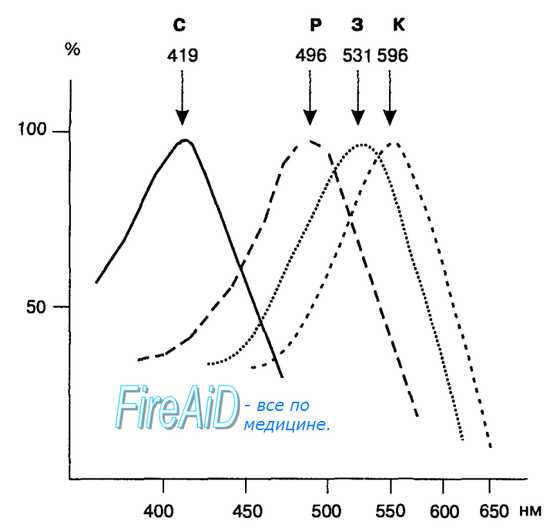
Рис. 17.7. Спектры поглощения четырех разновидностей фоторецепторов. Зрительный пигмент палочек родопсин (Р) имеет максимум поглощения световых волн длиною 496 нм, но способен также к поглощению коротких и длинных волн светового диапазона. Зрительный пигмент колбочек, чувствительных к синему цвету (С), имеет максимум поглощения 419 нм и не поглощает длинные волны оптического диапазона. Пигмент колбочек, чувствительных к зеленому цвету (3), имеет максимум поглощения при 531 нм, а пигмент чувствительных к красному цвету колбочек (К) максимально поглощает волны длиной 596 нм.
Родопсин наиболее чувствителен к электромагнитным волнам длиной около 500 нм, но хорошо поглощает и другие волны в диапазоне от 400 до 600 нм. Способность родопсина поглощать волны почти всего светового диапазона позволяет палочкам обеспечить только ахроматическое, т. е. черно-белое, зрение и лишает их возможности различать цвет. Высокое содержание зрительного пигмента в палочках и его способность суммировать фотоны, поглощенные в течение около 100 мс, делает палочки наиболее чувствительными фоторецепторами сетчатки. При слабом сумеречном освещении зрение обеспечивают только палочки, способные возбуждаться вследствие поглощения всего лишь одного фотона. Палочки образуют скотопическую систему, илисистему ночного зрения.
Зрительные пигменты колбочек подобны родопсину палочек и состоят из светопоглощающей молекулы ретиналя и опсина, который отличается составом аминокислот от белковой части родопсина. Кроме того, колбочки содержат меньшее, чем палочки, количество зрительного пигмента, и для их возбуждения требуется энергия нескольких сотен фотонов. Поэтому колбочки активируются лишь при дневном или достаточно ярком искусственном освещении, они образуют фотопическую систему, или систему дневного зрения.
В сетчатке человека существуют три типа колбочек, различающихся между собой по составу аминокислот в опсине зрительного пигмента. Различия в белковой части молекулы определяют особенности взаимодействия каждой из трех форм опсина с ретиналем и специфическую чувствительность к световым волнам разной длины (рис. 17.7). Колбочки одного из трех типов максимально поглощают короткие световые волны с длиной 419 нм, что необходимо для восприятия синего цвета. Другой тип зрительного пигмента наиболее чувствителен к волнам средней длины и имеет максимум поглощения при 531 нм, он служит для восприятия зеленого цвета. Третий тип зрительного пигмента максимально поглощает длинные волны с максимумом при 559 нм, что позволяет воспринимать красный цвет. Наличие трех типов колбочек обеспечивает человеку восприятие всей цветовой палитры, в которой существует свыше семи миллионов цветовых градаций, тогда как скотопическая система палочек позволяет различать лишь около пятисот черно-белых градаций.
Рецепторный потенциал палочек и колбочек. Ток ионов через мембрану фоторецептора в темноте и на свету.
Специфической особенностью фоторецепторов является темновой ток катионов через открытые мембранные каналы внешних сегментов (рис. 17.8). Эти каналы открываются при высокой концентрации циклического гуанозинмонофосфата, который является вторичным посредником рецепторного белка (зрительного пигмента). Темновой ток катионов деполяризует мембрану фоторецептора до приблизительно —40 мВ, что приводит к выделению медиатора в его синаптическом окончании. Активированные поглощением света молекулы зрительного пигмента стимулируют активность фосфодиэстеразы — фермента, расщепляющего цГМФ, поэтому при действии света на фоторецепторы в них уменьшается концентрация цГМФ. В результате управляемые этим посредником катионные каналы закрываются, и ток катионов в клетку прекращается.

Рис. 17.8. Ток ионов через мембрану фоторецептора в темноте и на свету. Ионы натрия поступают в фоторецепторы через катионные каналы внешнего сегмента, которые открываются при высокой концентрации цГМФ в темноте (темновой ток). Поступление в клетку ионов натрия вызывает деполяризацию мембраны фоторецептора до —40 мВ. На свету концентрация цГМФ уменьшается, что приводит к закрытию катионных каналов. Непрекращающийся выход ионов калия ведет к гиперполяризации мембраны, которая представляет собой рецепторный потенциал.
Вследствие непрерывного выхода ионов калия из клеток, мембрана фоторецепторов гиперполяризуется приблизительно до —70 мВ, эта гиперполяризация мембраны является рецепторным потенциалом. При возникновении рецепторного потенциала прекращается выделение глутамата в синаптических окончаниях фоторецептора.
Фоторецепторы образуют синапсы с биполярными клетками двух типов, различающихся по способу управления хемозависи-мыми натриевыми каналами в синапсах. Действие глутамата приводит к открытию каналов для ионов натрия и деполяризации мембраны одних биполярных клеток и к закрытию натриевых каналов и гиперполяризации биполярных клеток другого типа. Наличие двух типов биполярных клеток необходимо для формирования антагонизма между центром и периферией рецептивных полей ганглиозных клеток.
Адаптация фоторецепторов к изменениям освещенности. Световая адаптация. Десенситизация. Темновая адаптация.
Временное ослепление при быстром переходе от темноты к яркому освещению исчезает спустя несколько секунд благодаря процессусветовой адаптации. Одним из механизмов световой адаптации является рефлекторное сужение зрачков, другой зависит от концентрации ионов кальция в колбочках. При поглощении света в мембранах фоторецепторов закрываются катионные каналы, что прекращает вхождение ионов натрия и кальция и уменьшает их внутриклеточную концентрацию. Высокая концентрация ионов кальция в темноте подавляет активность гуанилатциклазы — фермента, определяющего образование цГМФ из гуанозинтрифосфата. Вследствие снижения концентрации кальция, обусловленного поглощением света, активность гуанилатциклазы повышается, что ведет к дополнительному синтезу цГМФ. Повышение концентрации этого вещества приводит к открытию катионных каналов, восстановлению тока катионов в клетку и, соответственно, способности колбочек отвечать на световые раздражители как обычно. Низкая концентрация ионов кальция способствует десенситизации колбочек, т. е. уменьшению их чувствительности к свету. Десенситизация обусловлена изменением свойств фосфодиэстеразы и белков катионных каналов, становящихся менее чувствительными к концентрации цГМФ.

Способность различать окружающие предметы исчезает на некоторое время при быстром переходе от яркого света к темноте. Она постепенно восстанавливается в ходе темновой адаптации, обусловленной расширением зрачков и переключением зрительного восприятия с фотопической системы на скотопическую. Темновую адаптацию палочек определяют медленные изменения функциональной активности белков, приводящие к повышению их чувствительности. В механизме темновой адаптации участвуют и горизонтальные клетки, способствующие увеличению центральной части рецептивных полей в условиях низкой освещенности.
Рецептивные поля клеток сетчатки. Прямой путь передачи сигналов от фоторецепторов к ганглиозной клетке. Непрямой путь передачи сигналов.
Выходные сигналы, передаваемые в ЦНС от сетчатки, возникают только в ганглиозных клетках, импульсная активность которых зависит от возбуждения фоторецепторов, а затем биполярных клеток, входящих в округлое рецептивное поле ганглиозной клетки. Размер рецептивных полей и количество фоторецепторов, относящихся к одному рецептивному полю, варьируют от минимального в области центральной ямки до наибольшего на периферии сетчатки. Малые рецептивные поля служат для различения мелких деталей наблюдаемых объектов в тех случаях, когда соседние детали воспринимаются под углом в несколько угловых минут. Большие рецептивные поля вмещают изображение целого объекта, воспринимаемого под углом в несколько угловых градусов (1гр соответствует рецептивному полю на поверхности сетчатки с диаметром около 0,25 мм).
Существуют два пути для передачи сигналов от фоторецепторов к ганглиозной клетке: прямой и непрямой.
Прямой путь начинается от фоторецепторов, расположенных в центре рецептивного поля и образующих синапс с биполярной клеткой, которая через другой синапс действует на ганг-лиозную клетку.
Непрямой путь берет начало от фоторецепторов периферии рецептивного поля, которая с центром состоит в реципрокных отношениях, обусловленных тормозным действием горизонтальных и амакрино-вых клеток (латеральное торможение).
Рецептивные поля с оn-центрами и off-центрами. On-нейроны. Off-нейроны. Ганглиозная клетка on-типа. Ганглиозная клетка off-типа.
В сетчатке человека имеются два типа ганглиозных клеток, отличающихся реакцией на точечные световые стимулы, воздействовавшие на центр или периферию их рецептивного поля (рис. 17.9). Примерно половина ганглиозных клетоквозбуждается действием света на центр рецептивного поля и тормозится при действии светового стимула на периферию рецептивного поля. Такие клетки принято называть оn-нейронами. Другая половина ганглиозных клеток возбуждается действием светового раздражителя на периферию рецептивного поля и тормозится в ответ на световую стимуляцию центра рецептивного поля — они получили название off-нейронов.

Рис. 17.9. Рецептивные поля ганглиозных клеток сетчатки с on- и off-центрами.
A. Рецептивное поле ганглиозной клетки образуют все фоторецепторы и биполярные клетки, имеющие с нею синаптические контакты. Ганглиозные клетки постоянно генерируют потенциалы действия, частота возникновения которых зависит от активности фоторецепторов и биполярных клеток, входящих в ее рецептивное поле.
Б. Ганглиозная клетка on-типа увеличивает частоту электрических разрядов в ответ на световое раздражение центра рецептивного поля и снижает свою электрическую активность при действии светового раздражителя на периферию рецептивного поля.
B. Ганглиозная клетка off-типа тормозится при действии света на центр ее рецептивного поля и увеличивает частоту нервных импульсов в ответ на раздражение периферии рецептивного поля.
Рецептивные поля ганглиозных клеток обоих типов в сетчатке представлены поровну, чередуясь друг с другом. Оба типа клеток очень слабо отвечают на равномерную диффузную засветку всего рецептивного поля, а наиболее сильным раздражителем для них является световой контраст, т. е. различная интенсивность засветки центра и периферии. Именно контрастирование деталей изображения дает необходимую информацию для зрительного восприятия в целом, тогда как абсолютная интенсивность отраженного от наблюдаемого объекта света не столь важна. Восприятие граней, т. е. восприятие контраста между соседними поверхностями с разной освещенностью, является наиболее информативным признаком изображения, определяющим протяженность и позиции разных объектов.
Рецептивные поля цветового восприятия. Восприятие цвета. Первичные цвета. Монохромазия. Дихромазия. Трихромазия.
Восприятие цвета основано на существовании шести первичных цветов, образующих три антагонистичные, или цветооппонентных, пары: красный — зеленый, синий — желтый, белый — черный. Ганглиозные клетки, передающие в центральную нервную систему информацию о цвете, различаются организацией своих рецептивных полей, состоящих из комбинаций трех существующих типов колбочек. Каждая колбочка предназначена для поглощения электромагнитных волн определенной длины, однако сами они не кодируют информацию о длине волны и способны реагировать на очень яркий белый свет. И только наличие в рецептивном поле ганглиозной клетки антагонистичных фоторецепторов создает нейронный канал для передачи информации об определенном цвете. При наличии только одного типа колбочек (монохромазия) человек не способен различить ни один цвет и воспринимает окружающий мир в черно-белой градации, как при скотопическом зрении. При наличии только двух типов колбочек (дихромазия) цветовое восприятие ограничено, и лишь существование трех типов колбочек (трихромазия) обеспечивает полноту цветового восприятия. Возникновение монохромазиии дихромазии у человека обусловлено генетическими дефектами Х-хромосомы.
Концентрические широкополосные ганглиозные клетки имеют округлые рецептивные поля on- или off-типа, которые образованы колбочками, но предназначены для фотопического черно-белого зрения. Белый свет, попадающий в центр или на периферию такого рецептивного поля, возбуждает или тормозит активность соответствующей ганглиозной клетки, которая в итоге передает информацию об освещенности. Концентрические широкополосные клетки суммируют сигналы от колбочек, поглощающих красный и зеленый цвет и расположенных в центре и на периферии рецептивного поля. Поступление сигналов от колбочек обоих типов происходит независимо друг от друга, а потому не создает цветового антагонизма и не позволяет широкополосным клеткам дифференцировать цвет (рис. 17.10).

Рис. 17.10. Рецептивные поля ганглиозных клеток, образованные колбочками трех типов.
A. Концентрические широкополосные клетки имеют в своем рецептивном поле колбочки для восприятия красного (К) и зеленого (3) цветов. Колбочки обоих типов расположены в рецептивном поле так, что их входные сигналы суммируются независимо друг от друга, поэтому широкополосные ганглиозные клетки не передают информацию о цвете, а реагируют лишь на световой контраст между центром и периферией рецептивного поля как on- или off-нейроны. Б. Простые противоцветные клетки двух разновидностей, отвечающие на стимуляцию зеленым цветом центра или периферии рецептивного поля (красный цвет действует как антагонист зеленому).
B. Простые противоцветные клетки двух разновидностей, различающиеся по характеру ответа на действие красного цвета в центре или на периферии рецептивного поля (зеленый цвет действует как антагонист красному).
Г. Колбочки, поглощающие короткие волны (синий цвет), находятся в антагонистических отношениях со входящими в общее рецептивное поле колбочками, поглощающими средние и длинные волны светового диапазона (зеленый и красный цвет).
Наиболее сильным раздражителем для концентрических противоцветных ганглиозных клеток сетчатки является действие антагонистических цветов на центр и периферию рецептивного поля. Одну разновидность противоцветных ганглиозных клетоквозбуждает действие красного цвета на центр ее рецептивного поля, в котором сосредоточены колбочки, чувствительные к красной части спектра, и зеленого цвета — на периферию, где имеются колбочки, чувствительные к нему. У другой разновидности концентрических противоцветных клеток в центре рецептивного поля расположены колбочки, чувствительные к зеленой части спектра, а на периферии — к красной. Эти две разновидности концентрических противоцветных клеток различаются ответными реакциями на действие красного или зеленого цвета на центр или периферию рецептивного поля подобно тому, как on- и off-нейроны различаются в зависимости от воздействия света на центр или периферию рецептивного поля. Каждая из двух разновидностей противоцветных клеток представляет собой нейронный канал, передающий информацию о действии красного или зеленого цвета, причем передача информации тормозится действием антагонистического или оппонентного цвета.
Оппонентные отношения при восприятии синего и желтого цветов обеспечиваются в результате объединения в рецептивном поле колбочек, поглощающих короткие волны (синий цвет) с комбинацией из колбочек, реагирующих на зеленый и красный цвет, что при смешении дает восприятие желтого цвета. Синий и желтый цвет оппонентны по отношению друг к другу, и сочетание в рецептивном поле колбочек, поглощающих эти цвета, позволяет противоцветной ганглиозной клетке передавать информацию о действии одного из них. Каким именно окажется этот нейронный канал, т. е. передающим информацию о синем или желтом цвете, определяет расположение колбочек внутри рецептивного поля концентрической противоцветной клетки. В зависимости от этого нейронный канал возбуждается синим или желтым цветом и тормозится оппонентным цветом.
М- и Р-типы ганглиозных клеток сетчатки. Магноцеллюлярные (М-клетки) клетки. Парвоцеллюлярные (Р-клетки) ганглиозные клетки сетчатки.
Зрительное восприятие происходит в результате согласования друг с другом различных сведений о наблюдаемых объектах. Но на низших иерархических уровнях зрительной системы, начиная с сетчатки глаза, осуществляется независимая переработка информации о форме и глубине объекта, о его цвете и его движении. Паралелльная переработка информации об этих качествах зрительных объектов обеспечивается специализацией ганглиозных клеток сетчатки, которые подразделяются на магноцеллюлярные (М-клетки) и парвоцеллюлярные (Р-клетки).
В большом рецептивном поле относительно крупных М-клеток, состоящем преимущественно из палочек, может проецироваться цельное изображение крупных объектов: М-клетки регистрируют грубые признаки таких объектов и их движение в зрительном поле, отвечая на раздражение всего рецептивного поля непродолжительной импульсной активностью. Клетки Р-типа имеют малые рецептивные поля, состоящие преимущественно из колбочек и предназначенные для восприятия мелких деталей формы объекта или для восприятия цвета. Среди ганглиозных клеток каждого типа имеются как on-нейроны, так и off-нейроны, дающие наиболее сильный ответ на раздражение центра или периферии рецептивного поля. Существование М- и Р-типов ганглиозных клеток позволяет разделить информацию о разных качествах наблюдаемого объекта, которая перерабатывается независимо в параллельных путях зрительной системы: о тонких деталях объекта и о его цвете (пути начинаются от соответствующих рецептивных полей клеток Р-типа) и о движении объектов в зрительном поле (путь от клеток М-типа).
Проводящие пути и центры зрительной системы. Зрительный нерв. Зрительные тракты. Глазодвигательный рефлекс.
Аксоны ганглиозных клеток образуют зрительный нерв, состоящий приблизительно из одного миллиона волокон. Войдя через зрительные отверстия в череп, правый и левый зрительные нервы сходятся друг с другом и частично перекрещиваются: волокна от носовых половин сетчаток переходят на противоположную сторону, волокна от височных половин сетчаток следуют ипсилатерально. В результате частичного перекреста образуются правый и левый зрительные тракты, передающие информацию от противоположной половины зрительного поля.

Окончания аксонов ганглиозных клеток на нейронах претектальной области среднего мозга служат для осуществлениязрачкового рефлекса. Аксоны, следующие к верхним буграм четверохолмия, переносят информацию, необходимую для осуществления глазодвигательных рефлексов и ориентировочных реакций. Проводящий путь от сетчатки к гипоталамусу предназначен для синхронизации эндогенного циркадианного 24-часового ритма с естественным чередованием светлого и темного времени суток. Собственно сенсорная функция, т. е. восприятие зрительной информации, обеспечивается при участии латерального коленчатого тела, принимающего информацию от сетчатки и передающего ее в зрительную кору.
Латеральное коленчатое тело. Функциональная организация латерального коленчатого тела. Рецептивные поля латерального коленчатого тела.
Аксоны ганглиозных клеток образуют топографически организованные соединения с нейронами латерального коленчатого тела, которые представлены шестью слоями клеток. Два первых слоя, расположенные вентрально, состоят из магноцеллюлярных клеток, имеющих синапсы с М-клетками сетчатки, причем первый слой получает сигналы от носовой половины сетчатки контралатерального глаза, а второй — от височной половины ипсилатерального глаза. Остальные четыре слоя клеток, расположенные дорсальнее, получают сигналы от Р-клеток сетчатки: четвертый и шестой — от носовой половины сетчатки контралатерального, а третий и пятый — от височной половины сетчатки ипсилатерального глаза. В результате такой организации афферентных входов в каждомлатеральном коленчатом теле, т. е. левом и правом, формируются шесть расположенных точно одна над другой нейронных карт противоположной половины зрительного поля. Нейронные карты организованы ретинотопически, в каждой из них около 25 % клеток получают информацию от фоторецепторов центральной ямки.

Рецептивные поля нейронов латерального коленчатого тела имеют округлую форму с центрами on- или off-типа и антагонистичной по отношению к центру периферией. К каждому нейрону конвергирует небольшое количество аксонов ганглиозных клеток, и потому характер передающейся зрительной коре информации здесь почти не изменяется. Сигналы от парвоцеллюлярных и магноцеллюлярных клеток сетчатки перерабатываются независимо друг от друга и передаются в зрительную кору параллельными путями. Нейроны латерального коленчатого тела получают от сетчатки не более 20 % афферентных входов, а остальные афференты образованы в основном нейронами ретикулярной формации и коры. Эти входы в латеральное коленчатое телорегулируют передачу сигналов от сетчатки к коре.
Переработка зрительной сенсорной информации в коре. Проекционная зрительная кора. Световая грань. Комплексные нейроны. Двойные противоцветные клетки..
Проекционная зрительная кора расположена в затылочных долях каждого полушария (поле 17 или VI) и получает организованные ретинотопически афферентные входы от соответствующего, т. е. левого или правого, латерального коленчатого тела. Каждый участок проекционной коры соответствует определенным точкам сетчатки, на которые проецируются изображения предметов, расположенных в противоположной полушарию половине зрительного поля. Проекция центральной ямки и ближайших к ней участков сетчатки занимает около половины первичной зрительной коры. Информация о форме наблюдаемых объектов, об их цвете и их движении поступает по параллельным путям в разные слои первичной зрительной коры. Объединение информации обо всех качествах наблюдаемых объектов происходит в процессе ее переработки во вторичной зрительной и ассоциативной коре.
Аксоны нескольких нейронов латерального коленчатого тела передают информацию входным звездчатым клеткам коры, отростки которых конвергируют к одному из простых пирамидных нейронов первичной зрительной коры. Поэтому рецептивное поле последнего имеет форму вытянутого в длину прямоугольника, составленного из нескольких округлых рецептивных полей нейронов предыдущего уровня (рис. 17.11). Центральная часть такого рецептивного поля представляет собой on- либо off-зону, антагонистичную по отношению к периферии. Наиболее эффективным раздражителем для простых нейронов является действующая вдоль их рецептивного поля световая грань (светлая полоса на темном фоне, темная линия на светлом фоне, контраст между темной и светлой плоскостью). В первичной зрительной коре имеется около 20 популяций простых нейронов, различающихся чувствительностью к определенному углу наклона световой грани.
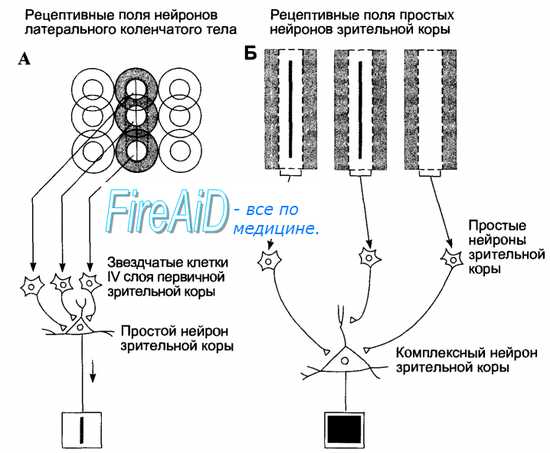
Рис. 17.11. Рецептивные поля нейронов разных иерархических уровней зрительной системы.
А. Нейроны латерального коленчатого тела имеют округлые рецептивные поля с on- или off-центрами, подобные рецептивным полям ганглиозных клеток. Конвергенция нейронов латерального коленчатого тела к простым нейронам проекционной зрительной коры (опосредованная звездчатыми клетками) формирует их рецептивные поля прямоугольной формы и максимальной чувствительности к определенному углу наклона линейного стимула.
Б. Конвергенция нейронов зрительной коры к комплексным нейронам создает рецептивное поле последних, в котором угол наклона линейных стимулов не имеет значения.
Наряду с простыми нейронами в первичной зрительной коре имеются комплексные нейроны, получающие афферентную информацию от нескольких простых нейронов и от входных клеток коры. Рецептивные поля комплексных нейронов больше, чем у простых нейронов, а линейная ориентация стимулов уже не имеет решающего значения для их возбуждения: самым значимым для них раздражителем служит движение грани или светового контура через рецептивное поле. Более половины комплексных нейронов глазодоминантны, т. е. сильнее реагируют на раздражение одного глаза, каждый из которых поставляет чуть отличающуюся информацию в связи с поперечной диспарацией. Глазодоминантные нейроны обеспечивают бинокулярное зрение, необходимое для определения пространственной глубины наблюдаемых объектов.
Ни простые, ни комплексные нейроны не реагируют на монотонные участки внутри объекта или фона — такие участки не дают информации, и восприятие внутренних участков зрительного объекта полностью определяется их границами.
Анализ информации о цвете в первичной зрительной коре осуществляют сосредоточенные там в виде так называемых капель (blobs)двойные противоцветные клетки, получающие афферентную информацию от простых противоцветных клеток сетчатки. Двойные противоцветные клетки возбуждаются (или тормозятся) при попадании одного цвета в центр рецептивного поля, а антагонистичного цвета — на периферию. Первичная зрительная кора не может обеспечить цветовое восприятие, которое осуществляется на следующей ступени преобразования поступившей информации в регионах V2 и V4 (последний расположен на стыке теменной и височной областей).
Зрительное восприятие. Магноцеллюлярный путь. Парвоцеллюлярный путь. Восприятие формы, цвета.
Целостное восприятие окружающего мира происходит в результате объединения всей информации, поступившей по параллельным путям зрительной системы. Магноцеллюлярный путь обеспечивает регистрацию движения объектов, восприятие пространственной глубины и определение пространственных отношений между объектами. В объединении этой информации помимо первичной зрительной коры участвуют регионы V2 (поле 18), V3 (поле 19), V5 или МТ (медиотемпоральный), MST (область коры, проходящая вдоль верхней височной борозды) и заднетеменной коры (поля 5 и 7). Парвоцеллюлярный путь, берущий начало от on- и off-ганглиозных клеток сетчатки и служащий для анализа информации о контурах объекта и линейной ориентации его деталей, после переработки в регионе VI продолжается в направлении нижних височных долей коры, при участии которых происходит восприятие формы наблюдаемых объектов.Параллельный парвоцеллюлярный путь начинается от концентрических противоцветных ганглиозных клеток сетчатки и предназначается для формирования зрительного представления о цвете объекта. Необходимая информация последовательно перерабатывается в регионах VI, V2 и V4, где происходит объединение зрительного восприятия формы и цвета. В регионе V4 расположены нейроны, воспринимающие определенный цвет, а не длину волн, соответствующих тому или иному цвету. С активностью этих нейронов связан феномен цветового постоянства, т. е. способность воспринимать цвет одних и тех же объектов неизменным, несмотря на естественные изменения освещенности на протяжении светового дня.

Восприятие образуется из элементарных зрительных ощущений подобно тому, как музыкальная мелодия складывается из отдельных звуков определенной высоты и узнается независимо от того, в какой тональности она сыграна. Процесс зрительного восприятия состоит из нескольких этапов. Первый из них осуществляется очень быстро и состоит в определении важнейших контуров объектов, их границ и текстуры поверхности, когда внимание еще не фиксировано на каких-либо деталях. После этого происходит фокусирование внимания на деталях, которые, в зависимости от их важности для целостного восприятия, могут быть просмотрены несколько раз, тогда как другие, менее информативные остаются незамеченными. Например, при рассматривании человеческого лица внимание смотрящего помимо общего контура более всего сосредоточено на таких деталях, как глаза и губы, тогда как щеки обычно привлекают внимание в меньшей мере. Зрительное восприятие не сводится к элементарному отражению зрительного поля, оно складывается и в результате конструктивной и высокопроизводительной активности всех участвующих в этом процессе регионов мозга: благодаря этому человек может с первого взгляда узнавать лица, вещи или пейзажи.
И палочки, и колбочки содержат вещества, которые распадаются под действием света, в результате возбуждаются нервные волокна, выходящие из глаза. Светочувствительное вещество в палочках называют родопсином; состав светочувствительных веществ в колбочках, называемых пигментами колбочек, или цветными пигментами, лишь немного отличается от родопсина. В этом разделе мы обсудим в основном фотохимию родопсина, но те же явления приложимы и к пигментам колбочек.
Наружный сегмент палочки, погруженный в пигментный слой сетчатки, примерно на 40% состоит из светочувствительного пигмента родопсина, или зрительного пурпура. Это вещество представляет собой соединение белка скотопсина и каротиноидного пигмента ретиналя (или ретинена). Важно, что ретиналь представлен в особой форме — 11 -цис-ретиналь, поскольку только эта цис-форма может связываться со скотопсином для синтеза родопсина. После поглощения световой энергии родопсин в течение незначительной доли секунды начинает распадаться.
Причиной этого является фотоактивация электронов в ретинальной части родопсина, что ведет к немедленному превращению цис-формы ретиналя в полностью-транс-форму, которая имеет ту же химическую структуру, что и цис-форма, но другую физическую структуру — прямую, а не изогнутую молекулу. Поскольку трехмерная ориентация реактивных участков полностью-транс-ретиналя больше не сходится с ориентацией реактивных участков белка скотопсина, эта форма ретиналя начинает отделяться от скотопсина. Непосредственный продукт реакции — батородопсин (прелюмиродопсин) — представляет собой частично расщепленную комбинацию полностью-транс-ретиналя и скотопсина.

Батородопсин — весьма нестабильное вещество, которое распадается в течение наносекунд до люмиродопсина. Последний, в свою очередь, распадается в течение микросекунд до метародопсина I, затем в течение примерно миллисекунды превращается в метародопсин II и, наконец, гораздо медленнее (в течение нескольких секунд) расщепляется на отдельные продукты — скотопсин и полностью-транс-ретиналь.
Именно метародопсин II, называемый также активированным родопсином, вызывает электрические изменения в палочках, которые затем передают зрительный образ в центральную нервную систему в форме потенциалов действия зрительного нерва, что будет изложено далее.
Восстановление родопсина. Первой стадией восстановления родопсина является обратное превращение полностью- транс-ретиналя в 11-цис-ретиналь. Этот процесс нуждается в метаболической энергии и катализируется ферментом ретиналь-изомеразой. Сразу после образования 11-цис-ретиналя он автоматически соединяется со скотопсином, вновь формируя родопсин, который остается стабильным, пока снова не начнется его распад при поглощении световой энергии.
Роль витамина А в формировании родопсина. На рисунке показан второй химический путь, с помощью которого полностью- транс-ретиналь может превращаться в 11-цис-ретиналь. Это происходит путем конверсии полностью-транс-ретиналя сначала в полностью-транс-ретинол — одну из форм витамина А. Затем под влиянием фермента изомеразы полностью- транс-ретинол превращается в 11-цис-ретинол. Наконец, 11-цис-ретинол конвертируется в 11-цис-ретиналь, который комбинируется со скотопсином, формируя новый родопсин.
Витамин А присутствует и в цитоплазме палочек, и в пигментном слое сетчатки. Следовательно, в норме при необходимости он всегда доступен для формирования нового ретиналя. С другой стороны, при избытке ретиналя в сетчатке он легко превращается снова в витамин А, уменьшая таким образом количество светочувствительного пигмента. Позднее мы увидим, что взаимопревращения ретиналя и витамина А особенно важны при долговременной адаптации сетчатки к различной интенсивности света.