

Генетическая регуляция биоритмов
Одной из характерных особенностей поведения различных животных является феномен периодичности, проявляющийся в том, что периоды активности чередуются у них с периодами покоя. Известно, например, что лев бодрствует 3—4 часа в сутки, а остальное время спит. Такого рода ритмы принято называть циркадными (от латинских слов "circa", что означает "вокруг" и "dies", что означает "день"). В некоторых случаях животные могут ориентироваться во времени суток благодаря способности, называемой временной памятью. Например, дрессируя пчел, можно добиться того, что они будут искать корм в определенное время дня. Если в течение нескольких дней давать пчелам корм в одно и то же время, они будут продолжать искать его в это же время даже после прекращения такого кормления. Иными словами, животные обладают своеобразными биологическими часами, которые подобно нашим современным часам связаны с циклическими процессами и могут быть независимыми от чередования дня и ночи. Отдельные случаи эндогенной циклической активности были описаны у животных еще в 1894 г., когда А. Кизель (A. Kiesel) обнаружил, что миграция пигмента у членистоногих продолжается и в отсутствие чередования света и темноты.
У млекопитающих выявлен ритм колебаний содержания гликогена в печени, у насекомых суточная периодичность вылета имаго из куколок, у раков - суточные колебания окраски, у человека - способность просыпаться точно в определенное время или в течение дня точно соблюдать определенные сроки, не пользуясь внешними указателями времени. Проводились поиски тех центров, которые "управляют" процессами циклической активности. У тараканов, например, таковыми оказались нейросекреторные клетки головного мозга. Нейральная природа


SO
55
50
во -
BPr,
~ ЬОсек
;ЛЛААЛЛАА
Д икий
тип
икий
тип
so
BO 55 50
I
-УОсек
Время
я![]() меет
молекулярный вес около
127 000. Две минорных РНК
продуцируются посредством
альтернативного сплай-янга
близ З1
- конца, так что юзможно
образование трех елков,
однако значение их
меет
молекулярный вес около
127 000. Две минорных РНК
продуцируются посредством
альтернативного сплай-янга
близ З1
- конца, так что юзможно
образование трех елков,
однако значение их
еизвестно. Продукт per локуса образуется во многих тканях, включая ибриональную, куколочную и взрослую нервную систему, пищевод, шки, яичники. Этот продукт локализуется преимущественно в кле-)чных ядрах и, возможно, действует как транскрипционный фактор, н сходен с белками, кодируемыми тремя другими генами, кодирующи-[ (а)человеческий aryl hydrocarbon рецептор, ядерный транспортер ого рецептора и singleminded белок Drosophila melanogaster. Эти гены цержат домен размером примерно в 270 аминокислот, который ис-пьзуется для димеризации и взаимодействия с партнером, который (ержит ДНК-связывающую область и потому функционирует как шскрипционный регулятор.
Известны и другие гены, оказывающие влияние на биоритм - это щепленные Clock и Andante локусы, слабо действующие в направле-[ укорочения или удлинения цикла соответственно. Белок PER, отно-щйся к протеогликанам, экспрессируется в мозге, в глиальных клет-и в некоторых нервных клетках. Экспрессируется он и в других тка-. Были предприняты попытки обнаружить клетки, ответственные за троль циркадного ритма. В результате в мозге был найден комплекс тральных нейронов, обнаруживающих циркадные колебания в со-кании PER белка. Оказалось, что синтез PER белка и per мРНК ха-геризуется ритмичностью, у мутантов per0 эта ритмичность наруша-. Найдено, что белок PER взаимодействует с другими белками, в ре-тате чего тормозится синтез per мРНК, и уровень содержания этой I снижается ночью, так что синтезируется меньше соответствующе-елка. У мух дикого типа цикл репрессии-дерепрессии синтеза per '. подчинен 24-часовому ритму. Если в геном мух per0 ввести индуци-[ую тепловым шоком копию гена рег+, обозначенную hspcper, то сгенные животные перестают проявлять аритмичность циркадного а и переходят в своем поведении на 24-часовой цикл активности. Z помощью иммуногистохимической реакции были показаны соот-^вующие циркадному ритму колебания в содержании PER белка в сых нейронах дрозофилы и в мелких нейронах мозга, которые тру-
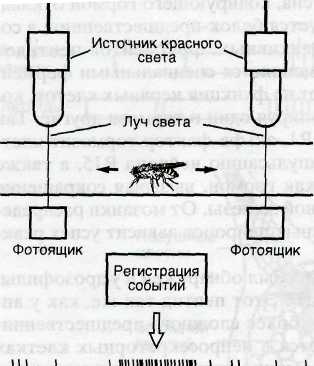
Рис. 7.9. Способ регистрации ] ного ритма у дрозофилы. Объяснение! в тексте. По Snustad et al., 1995.
Время дня
контроля биоритмов, возможно,! связанная с активностью опреде-1 ленных групп нейронов и нейросе-креторных клеток мозга, присуща, по-видимому, и другим животным. У млекопитающих, например, показано, что центром, упра-1 вляющим биоритмами, является I супрахиазматическое ядро гипоталамуса - отдела мозга, особенно богатого нейросекреторными | клетками.
Такая эндогенная ритмическая I
активность имеет наследственную основу. Особенно большие успехи были достигнуты в этом направлении при изучении биоритмов у дрозофилы, у которой открыт соответствующий ген period (per), локализованный в X хромосоме, в области ЗВ1-2 и контролирующий периодичность активности мухи в течение суток. Для измерения этой активности муху помещают в стеклянную трубку, каждый конец которой освещается красным светом. Муха не может видеть красный цвет. Пересечение ею луча света с каждой стороны трубки регистрируется и характеризует уровень активности животного (рис. 7.9). В период большей активности муха чаще пересекает луч света.
Известны мутанты щшофтаы с таменением характера циклической активности. В частности, per0 аллель характеризуется аритмичностью поведения, pers (short) аллель детерминирует цикличность с 19-часовым интервалом вместо "нормального" 24-часового, per1 (long) изменяет интервал "дикого типа" на 29-часовой. Локус per контролирует также время вылета имаго из куколок. Мухи per* вылетают из куколок утром, когда наиболее благоприятны условия для того, чтобы расправить и высушить крылья и отправиться в полет. Паттерн вылета синхронизирован в соответствии с 24-часовым циклом. Мутанты per0 не обнаруживают в этом отношении цикличности и могут вылететь в любое время дня и ночи. Мутанты с удлиненным и укороченным циклом имеют и соответственно измененные паттерны вылета. Интересно, что та-1 кой элемент ухаживания дрозофилы, как песня, также зависит от гена per (рис. 7.10).
Естественно, встает вопрос о молекулярно-генетических механизмах регуляции циркадного ритма. Ген per дрозофилы был изолирован и идентифицирован его белковый продукт PER. Тек per имеет 7 кб длины, 8 экзонов, первый из которых является некодирующим и синтезирует транскрипт длиной 4,5 кб. При этом возникает по крайней мере три различных РНК. Локус per был секвенирован, его продукт-полипептид
! нейроны активируются продуктами гена, кодирующего гормон откладки яиц (ГОЯ), причем сначала образуется белок-предшественник, в состав которого входит несколько активных фрагментов-пептидов (рис. 7.8). Этот предшественник расщепляется специальными ферментами, и продукты расщепления влияют на функции нервных клеток, которые управляют кладкой яиц, стимулируя одни и тормозя другие. Так бета-пептид возбуждает клетки L1 и R1, альфа-фактор тормозит клетки L2, L3, L4, L6, ГОЯ усиливает импульсацию нейрона R15, а также выделяется в кровоток и действует как гормон, вызывая сокращение мускулатуры в протоке гермафродитной железы. От мозаики распределения активированных и заторможенных нейронов зависит успех реак-| ции в целом.
Гомологичный с моллюском пептид был обнаружен и у дрозофилы. Это Phe-Met-Arg-Phe-NH2 (FMRF-amide). Этот пептид так же, как у ап-лизии, первоначально входит в состав более сложного предшественника. Этот предшественник синтезируется в нейросекреторных клетках мозга и, как и у аплизии, регулирует реакцию откладывания яиц, демонстрируя достаточно универсальный характер описанного механизма.