

Проблема прабатьківщина людства
Вступ
Розділ 1. Ступінь наукової розробки теми та джерельна база.
Історіографія проблеми.
Джерельна база дослідження.
Розділ 2. Африканська концепція прабатьківщини людства.
Розділ 3. Мультирегіональна теорія.
Розділ 4. Позатропічна прабатьківщина людства.
Висновки.
Список використаних джерел та літератури.
Перші ж представники Homo sapiens, які є нащадками Homo erectus, з'явилися понад 500 тисяч років тому і відрізнялися за статурою від людини сучасного типу. Їхні кістки були виявлені і в Африці, і в Європі, і в Азії. Тому виникає питання: а де ж виникла сучасна людина? В даний час існують такі основні гіпотези: африканська концепція прабатьківщини людства, мультирегіональна теорія та позатропічна прабатьківщина людства.
Розділ 1. Ступінь наукової розробки теми та джерельна база.
Історія вивчення австралопітеків веде свій початок з 1924 року, з знахідки черепа дитинчати гоминоида 3-5 років у Південно-Східному Трансваалі (нині ПАР) поблизу Таунг ("бебі з Таунг"). Викопний гоминоида отримав найменування африканського австралопітека - Avstralopitecus africanus Dагt, 1925 (від "avstralis"-південний). У наступні роки були відкриті й інші місцезнаходження австралопітеків Південної Африки-в Стеркфонтейн, Макапансгате, Сварт-Кранс, Кромдраае. Їх залишки виявлялися зазвичай в печерах: вони залягали в травертину-відкладеннях вуглекислих джерел, що випливають з вапняків, або безпосередньо в породах доломітового товщі. Спочатку нові знахідки отримували самостійні родові позначення-плезиантропов (Plesianthropus), парантропов (Paranthropus), але, за сучасними уявленнями, серед південноафриканських австралопітеків виділяється тільки один рід Avstralopithecus з двома видами: більш давнім ("класичним") грацильний австралопітеком (А. africanus) і більш пізнім масивним, або парантропов (A. robustus).
У 1959 р. австралопітеки були виявлені і в Східній Африці. Перша знахідка зроблена подружжям М. і Л. Лики в найдавнішому 1 шарі (пачці) Олдувайского ущелини на околиці плато Серенгеті в Танзанії; цей гоминоида, представлений досить тероморфним черепом з гребенями, отримав найменування "зинджантропа бойсова", тобто східноафриканського людини, оскільки в безпосередній близькості до нього були відкриті і кам'яні артефакти (Zinjanthropus boisei Leakey). Надалі залишки австралопітеків знайдено в ряді місць Східної Африки, зосереджених головним чином в області Східноафриканського рифта (рис. 19). Основні локуси знахідок пліоценових і пліоплейстоценових гоминоида у Східній Африці: Олдувай, Летолі, Гарус, Пеніндж (Танзанія); Кооби Фора, Ілерет, Канапоі, Лотагам, Лукейно, Чесованджа, Нгорора, Чемерон, Зап. Туркана (Кенія); Хадар, Омо, Бодо (Ефіопія). Зазвичай вони є більш-менш відкриті стоянки, що включають ділянки трав'янистої лісостепу. Серед восточноафриканских австралопітеків, мабуть, також зустрічаються більш ранні грацильний і пізніші масивні (або навіть "надмасивні") форми. Їх відносять відповідно до видів А. afarensis і (або) А. africanus і А. Bоisei (тип зінджанропа). Однак деякі сторони таксономії австралопітекових все ще залишаються предметом дискусій.
До нас тоящему часу з територій Південної і Східної Африки відомі вже залишки не менш ніж 500 індивідів. Австралопітеки, мабуть, могли зустрічатися і в інших регіонах Старого Світу: наприклад, так званий гігантопітека з Біласпур в Індії або яванська мегантроп до деякої міри нагадують масивних африканських австралопітеків. Втім, положення цих форм у таксономії і філогенії гоминоида не цілком ясно. Таким чином, хоча не можна виключити дифузію австралопітеків в південні райони Євразії, основна їх маса тісно пов'язана у своєму розселенні з африканським континентом, де вони зустрічаються на півдні до Хадара на північному сході Африки.
Головна частина знахідок восточноафриканских австралопітеків датується періодом від 4 до 1 млн. років тому, проте найдавніші двоногі, мабуть, з'явилися тут ще раніше-5 ,5-4, 5 млн років тому. У Південній Африці, де відсутні прямі калій-аргонові датування та оцінка віку знахідок проводиться за непрямими даними, встановлюються близькі хронологічні межі існування австралопітеків: 3-1 млн років тому.
Фізичний тип австралопітеків можна розглядати як наочну ілюстрацію мозаїчності еволюції на ранніх етапах розвитку людської лінії. Це були двоногі форми (біпеди) з малим мозком і дуже великими зубами з товстої емаллю (мегадонтія). Прямоходіння є найдавнішою системою гоминизации, що сформувалася вже 4-3,5 млн років тому, а можливо, й раніше. Відомо, що примати взагалі мають здатність утримувати тулуб у більш-менш випрямленій становищі, однак біпедія людини принципово інша, ніж в інших великих гоминоида, що приймають за часами випрямлена положення. Кращим індикатором прямоходіння на скелеті приматів є тазовий пояс. Основну роль у його перетворенні у предків людини відіграло расширени е клубової кістки допереду і одночасне посилення її середній частині, а також зміцнення крижово-клубових і тазостегнових суглобів і їх зближення; прогресивний розвиток елементів Муска ульно-зв'язкового апарату, фіксують розгинання ноги в тазостегновому і колінному суглобах. Відсутність подібної структури тазу у інших приматів спонукає їх пересуватися на двох ногах швидко, згинаючи тулуб вперед. Експериментально показано, що з енергетичних витрат двоногий хода людини при ходьбі з нормальною швидкістю ефективніше, ніж типова чотиринога хода ссавців або чим двоногий або чотиринога локомоция шимпанзе. Отже, біпедія давала предкам людини певні енергетичні переваги.
Австралопітеки в цілому мали вже цілком людський тип разгибательного апарату кульшового суглоба. Навіть у внутрішній структурі тазу, вивченої рентгенографічно, відзначено велику схожість між австралопітеків і людиною. Таким чином, австралопітеки мали вже постійної двоногій ходою і цим принципово відрізнялися від усіх відомих мавп, як сучасних, так і викопних.
Двоноге ходіння австралопітеків, мабуть, полегшувалося їх ма-лорослостью. Індивідуальні значення довжини тіла здебільшого потрапляють у межі 100-156 см; чітко виражений статевий диморфізм. Можливо, що малі розміри тіла мали адаптивне значення в умовах теплового перевантаження або недостатньої калорійності їжі.
Мозок австралопітеків за абсолютним розвитку знаходився в межах варіацій його маси у сучасних понгид. Обсяг мозку коливався індивідуально від 300 до 570 см куб. середні для грацильний і масивних австралопітеків становлять відповідно 440-450 см куб. і 516-518 см куб. Ступінь церебралізаціі у викопних приматів може бути визначена, зрозуміло, лише дуже приблизно. Розраховані на основі емпіричної формули Дюбуа (1914) Е = k * P ^ 0.56, де Е і Р-відповідно маса мозку і тіла, а "k"-коефіцієнт церебралізаціі, значення "k" варіюють у австралопітеків в межах 1,0-1 , 5 т. е. в середньому вони в 2-2,5 рази менше, ніж у сучасної людини (2,6-2,7), але трохи вище, ніж у високорозвинених сучасних понгид. Близькі результати виходять і при використанні рівняння регресії.
Найдавніші представники роду австралопітеків відомі зі Східної Африки. Це вид австралопітека афарського (А.afarensis Таiеb), названий так за основним місцезнаходженням в області афарского депресії-низинного району на півночі Ефіопії. У цьому регіоні протягом 1973-1976 рр.. франко-американською експедицією зроблено ряд унікальних відкриттів. Особливе значення мала знахідка в листопаді 1974 р. мініатюрного скелета дорослої жіночої особини, названої дослідниками "Люсі" (номер за каталогом АL-288-1). Збереглося близько 40% всіх кісток скелета, що вперше дозволило реконструювати риси зовнішнього вигляду найдавнішого австралопітека.
Зараз до виду афарского австралопітека зараховують ранніх австралопітеків з місцезнаходжень Хадар (Афар) і, мабуть, Омо і Середній Аваш в Ефіопії; Летолі і Гарус в Танзанії, можливо, Канапоі, Чемерон і деякі інші найдавніші знахідки в Кенії. Орієнтовна старовину цього виду: від 2,9 до 3,9, в окремих випадках-до 4-5 млн років тому (Канапоі, Чемерон). Якщо погодитися з віднесенням всіх цих копалин до одного виду, він повинен був характеризуватися дуже високою гетерогенністю і винятково сильним статевим диморфізму, який міг бути наслідком значних відмінностей у темпах дозрівання самців і самок, як, наприклад, у сучасних горил, або ж статевого відбору. У цьому випадку висока морфологічна мінливість афарского австралопітеків тлумачиться як результат нерівномірного зростання (аллометріі) різних частин тіла. Крайня "об'едінітельская" позиція навіть знижує таксонів-мічного ранг афарского австралопітеків до підвидового, розглядаючи їх як підвид грацильний південноафриканських австралопітеків-А. Africanus tanzaniensis. Альтернативне таксономічне рішення: належність пліоценових двоногих до двох різних видів або навіть пологах, як спочатку вважали і самі дослідники хадарского місцезнаходження.
Розділ 2. Африканська концепція прабатьківщини людства.
Історія завжди непокоїть людину, а гіпотетичність переходить у наукову впевненість, коли історію досліджують на рівні ДНК. Невидиме для ока таким чином оповідає про минуле за його рідкісними рештками. Шведські вчені дійшли висновку, що наші загальні предки прийшли з Африки приблизно 50 000 років тому. Вони зібрали докази теорії про те, що саме того періоду і саме на тому континенті з’явилася сучасна людина homo sapiens і почала перебиратися до інших земель. Такий висновок дослідники університету міста Упсала зробили після ретельного вивчення фрагментів ДНК первісної людини. Дискусії про те, чи сучасна людина походить із Африки, а чи різні типи людини виникли в різночассі і в різних місцях нашої ойкумени, тривають ось уже близько 20 років. Дослідники цього шведського університету вивчили гени 53 жінок, чиї закам’янілі рештки було знайдено в різних закапелках світу. Учені зосередилися на аналізі особливого виду ДНК (мітохондріальна ДНК), яка передається лише від матері до дитини і змінюється певним та передбачуваним способом із покоління до покоління. Завдяки таким дослідам було зроблено висновок, що всі 53 жінки мають однакові генетичні корені і походять від спільних африканських предків. Водночас, як переконані тепер учені, приблизно 50 тисяч років тому почали відбуватися зміни в азіатській та європейській лініях людського виду. Подібні дослідження ДНК проводилися і в минулому, але саме шведи подолали технічні проблеми, а відтак провели значно ретельніший, аніж у минулому, аналіз цього виду ДНК. Коментуючи результати дослідження в науковому журналі Nature, незалежний спеціаліст з університету Пенсильванії в США сказав, що шведським ученим вдалося надати переконливі докази того, що homo sapiens уперше з’явився саме в Африці.
Зараз більшість вчених вважають колискою людства Африку. З трьох видів приматів, останки яких знайдені в Єгипті (оазис Файюм), що жили там близько 30 млн. років тому: арідіум (apidium), олігопітек (oligopithtcus) і парапітек (parapithecus), - родовід первісних людей починають з парапітека (грец. para - біля, близько; pithecos - мавпа). Розвиток його найближчого нащадка - пропліопітека (propliopithecus, грец. pro - раніше, попереду; pithecos - мавпа), привів до поділу приматів - гоміноїдів на дві сім'ї: понгідових (людиноподібних) - гігантопітек (Gigantopithecus), пліопітек (pliopithecus) і сівапітек (sivapithecus ), і гомінінів (предків людини). Від пліопітеків, перших мавп що залишили Африку, походять сучасні гібони (відділилися від загального стовбура гомінідів близько 18 млн. років тому), а від сівапітеків, що мешкали в Азії близько 12-10 млн років тому [Рамапітека (ramapithecus), що раніше вважався однією з ланок еволюції людини, зараз визнають самкою сівапітека.] - орангутанги.
Розвиток гомінінів привів до виникнення дріопітеків (грец. drio - гущавина, дрімучий ліс, pithecos - мавпа, тобто буквально: «лісові мавпи»). Вони мешкали в Африці, Європі та Азії, але за своїм складом не були однорідні. І близько 7 млн. років тому розділилися на дві гілки: панід, які є предками сучасних горил і шимпанзе, і гомінід (предків людини). Якщо дріопітеки пересувалися на чотирьох кінцівках, то їх нащадки гомініди перейшли до прямоходіння. Самими древніми видами гомінідів зараз вважаються Sahelanthropus tchadensis (7 млн. років тому) і Orrorin tugenensis (6 млн. років тому). За ними йдуть Ardipithecus (6,3 - 3,9 млн. років тому), з видами Ar. kadabba і Ar. Ramidus (Мал.1).
У вересні 2009 року було опубліковано повідомлення про знахідку найстарішого відомого предка людини, вік якого становить 4,4 млн. років тому. Виявлений під час розкопок у центральній частині національного парку Аваш в Ефіопії вид отримав назву Ardipithecus ramidus. Його представники були прямоходячими, а реконструкція скелета особини жіночої статі, що отримала ім'я Арді, показала, що важила вона близько 50 кілограм при зростанні 1,2 метра.
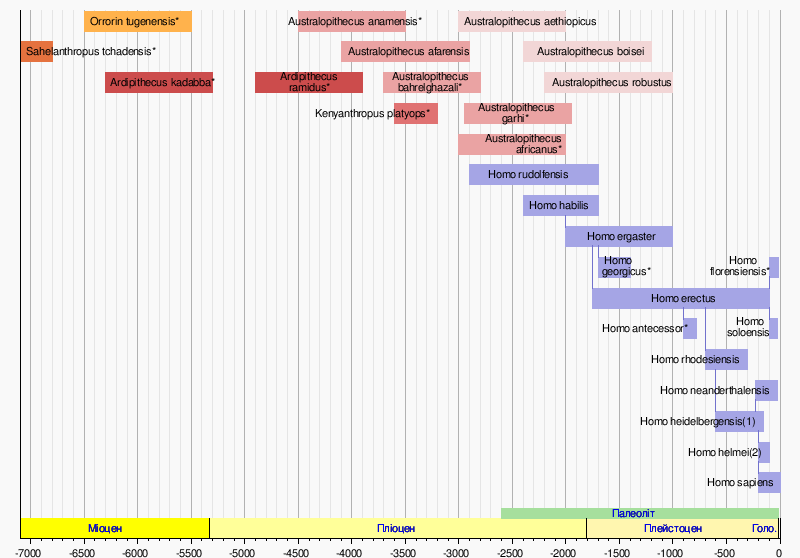
Мал. 1. Еволюція людини, діаграма видів.
Вік видів позначених * подано приблизно. - Вертикальні відрізки між видами роду Homo показують ймовірні розділення (гіпотеза "розщеплювача" ("splitter"), єдиного предка).
(1) або Homo sapiens архаїчна стара; (2) або Homo sapiens архаїчна остання.
Подальша еволюція Ardipithecus ramidus призвела до появи - австралопітеків (australopithecus, від латинського слова australis - південний, грец. pithecos - мавпа), близько 4 млн. років тому. Вони почали вживати природні предмети - палиці, роги, кістки, камені, - як знаряддя праці. Австралопітеки жили тільки на африканському континенті, в час від 4,5 до 1 млн. років тому. Відомо їх декілька видів: Australopithecus anamensis, Australopithecus afarensis (афарнский), Australopithecus bahrelghazali, Australopithecus aethiopiocus, Australopithecus africanus (граціальний, карликовий), Australopithecus garhi, Australopithecus boisei, Australopithecus robustus (великий), Australopithecus sediba.
У період між 3,0 - 1,5 млн. років тому стався стійкий поділ між австралопітеками і родом Homo (Людина). Предком сучасної людини був афарский австралопітек (A. afarensis), що жив 4,1 - 2,9 млн. років тому. До цього виду належить знаменита Люсі, знайдена в 1974 р. в Ефіопії, і яка жила близько 3,2 млн. років тому. А знахідка в 2010 р. "Kadanuumuu", або мовою афар "Великої людини", вік котрого 3,6 млн. років, довела, що A. afarensis були прямоходячим. У результаті подальшої еволюції, близько 2,4 млн. років тому, у Східній Африці, виділяється, морфологічно належний ще до австралопітеків вид Homo habilis («людина уміла»), який не лише використовував, але і почав виготовляти знаряддя праці.
Дату появи «людини умілої», виробника знарядь праці, приймають за початок періоду нижнього палеоліту (раннього давньокам'яного віку) - (близько 2 млн. років тому - 40 000 років до нашої ери, іноді виділяють середній палеоліт: 250/200 тис. років тому - 40 000 років до нашої ери), хоча перші знаряддя праці датуються віком близько 2,6 млн. років тому. [Слід мати на увазі, що періодизація кам'яного віку умовна, і місцеві відхилення вельми істотні.] Homo habilis створив найдавнішу культуру нижнього палеоліту - Олдувайську (галечну - застаріла назва) - (близько 2 - 1 млн. років тому). Її основні центри відкриті в східній Африці.
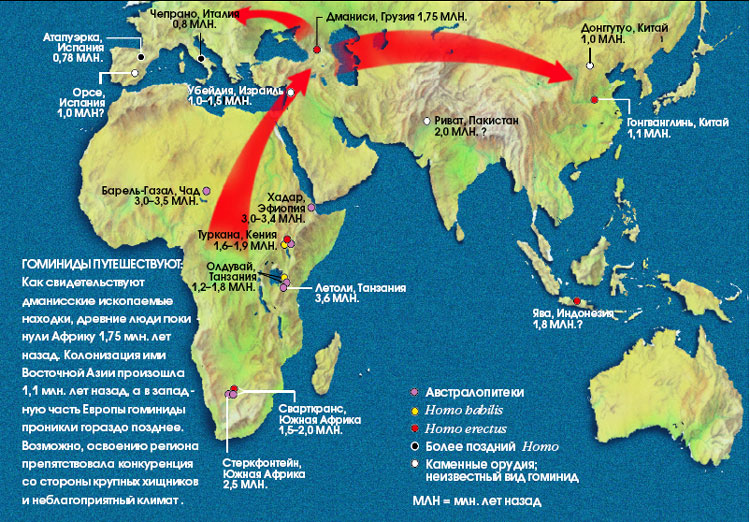
На зміну «людині умілій», яка був радше людиноподібної мавпою, близько 1,9 млн. років тому (в Африці) приходить вже первісна людина - архантроп (Homo erectus - «людина прямоходяча»). Він був вищий і важчій за «людину умілу», володів членороздільної мовою, регулярно використовував кам'яні знаряддя праці у своїй діяльності. Архантропи почав розселяться в Азію та Європу і близько 1 млн. років тому заселив всі три континенти. Але виявлені нещодавно викопні рештки підтримують гіпотезу про те, що, можливо, Homo erectus («людина прямоходяча») походить не від Homo habilis ("людини умілої»). У Кенії близько озера Туркана дослідники знайшли щелепу H. habilis віком 1,44 млн. років. У ті ж часи по цій землі ходили і представники H. erectus. Якщо два древніх виду гомінідів жили тут одночасно, то, ймовірно, вони незалежно походять від спільного предка. Найдавніші останки H. habilis і H. erectus, виявлені в Східній Африці, датуються 1,9 млн. років, і це говорить про те, що вони співіснували в даному регіоні протягом півмільйона років, а їх загальний прабатько повинен був жити 2-3 млн років тому.
Як би там не було, сам вид Homo erectus підрозділяється на ряд антропологічних типів: пітекантроп (pithicantropus, буквально «мавполюдина»), що жив з часу близько 700 тис. років тому на о. Ява і мав австралоідні риси; синантроп (sinanthropus - «китайська людина», або Homo pikinensis - «людина пекінська»), вік якого складає близько 400 тис. років тому, мав монголоїдні риси; атлантроп (від гір Атлас і грец. anthropos - чоловік), що жив з часу близько 360 тис. років тому на африканському узбережжі Середземного моря на території Алжиру і Марокко; олдувайской людина, яка жила в Сх. Африці; гейдельбергська людина (Homo heidelbergensis), що заселила Європу та Західну Азію близько 1 млн. років тому; близький останній за фізичним виглядом - афрікантроп, що заселяв Африку та ін. Найбільш пізнім знахідкам - 250 тис. років.
З «людиною прямоходячою» пов'язують виникнення ашельської культури нижнього палеоліту - (в Африці її вік складає близько 1,5 млн. років тому - 60 тис. років тому), яка поширювалася на величезні простори південної Азії, півдня Європи, і Африки аж до її крайніх південних областей. Тривалість ашельської культури в різних районах суттєво відрізняється, а деякі дослідники виділяють як попередницю ашельської шельську культуру. Ашельська культура принесла освоєння архантропом вогню, що прискорило його еволюційний розвиток.
Необхідно відзначити, що виявлені нещодавно в Грузії, біля міста Дманісі, викопні рештки гомінід свідчать про те, що найдавніші люди почали залишати Африку незабаром після того, як 1,9 млн. років тому на Землі з'явився Homo erectus. Однак питання про те, куди лежав шлях давніх мандрівників, залишається загадкою. Вік найстародавніших (після Дманісі) місць знаходження викопних решток гомінід в Азії складає трохи більше 1 млн. років, а в Європі - 800 тис. років (приналежність гоминидам скам'янілих кісток, виявлених у ряді місць на о. Ява віком 1,8 млн. років, піддається сьогодні сумнівам). Завдяки своїм анатомічним ознаками дманісська людина цілком може бути предком більш пізнього азіатського Н. erectus. Не виключено, однак, що ці гомініди виявилися "глухим кутом" людської еволюції - свого роду "гребенем хвилі", що вихлюпнулася з Африки лише на незначну частину території Євразії. Вчені вважають, що хвилі еміграції первісних людей з Африки піднімалися не раз.
Розділ 3. Мультирегіональна теорія
Мультирегіональних гіпотеза. Людина сучасного анатомічного типу виник незалежно на різних континентах з різних гілок Homo erectus. Однаковість людей підтримувалося системою міграцій (за: Stringer, 1989).
Рис. 4. Монофілетична гіпотеза (гіпотеза африканського походження людини). Людина сучасного типу виник в Африці і поширився звідти на всі континенти, витісняючи інших представників роду Homo (за: Stringer, 1989).
Згідно мультирегіональних гіпотези (рис. 3), люди різних рас мають різне походження: кожна виникла на своєму континенті від окремого представника виду Homo erectus. При цьому предкової раси складали єдину популяційної систему і, мігруючи, обмінювалися генами. Монофілетична ж гіпотеза стверджує, що всі люди мають спільне походження, і Homo sapiens, як і Homo erectus, виник в Африці, а потім розселився звідти по всіх континентах.
Прихильники обох гіпотез не сумніваються в походження людини від Homo erectus, але розходяться в питанні про час його виникнення. Згідно мультирегіональних гіпотезою, вік сучасної людини більше мільйона років, а за монофілетичної версії -- близько 500 тис. Однак ні антропологи, ні археологи не змогли поставити крапку в цьому суперечці. Корінний перелом у вирішенні проблеми виникнення людини, її розселення по континентах і виникнення рас внесли дослідження ДНК.
ДНК і відновлення історії людства
Історію популяцій людини можна простежити, порівнюючи представників різних рас і народів за тими фрагментами ДНК, за якими відмінності між індивідами набагато більше виражені, ніж у середньому по геному. Такі фрагменти називають ДНК-маркерами. Знаючи швидкість, з якою відбуваються мутації, і визначаючи кількісну міру відмінностей між геномом людей з різних популяцій, обчислюють час їх відділення від загальної предкової лінії. Просуваючись назад від геному сучасної людини, можна побудувати Філогенетичне древо Homo sapiens. Воно дозволяє робити певні висновки про генетичної історії людства.
Рис. 5. Фрагмент ДНК. Різні індивіди можуть відрізнятися один від одного по складу нуклеотидів (при постійній довжині фрагмента) або по довжині фрагмента (за рахунок вставки або випадання нуклеотидів).
Для дослідження використовують різні типи ДНК-маркерів. Слід зазначити ДНК мітохондрій (мтДНК) та Y-хромосоми (Y-ДНК), оскільки вони дозволяють простежити генетичну історію людства окремо по жіночій і чоловічій лініях. Перша передається тільки по материнській лінії, тому що мітохондрії знаходяться в цитоплазмі клітини, а цитоплазма нащадка (зиготи) визначається цитоплазмою яйцеклітини. Якщо дві людини мають спільного предка жіночої статі, то різниці між їх мтДНК покажуть, скільки поколінь відокремлює їх від жила століття або тисячоліття тому загальної прабабусі. Вивчення Y-ДНК дозволяє простежити еволюційні траєкторії по батьківській лінії, оскільки Y-хромосома передається тільки по чоловічій лінії. Вивчення решті, аутосомно частини геному, яка зосереджена в хромосомах, успадковується по обох лініях і в якій представлена велика частина генома людини, також надзвичайно важливо, бо дає маркери для вивчення комбінатівной мінливості, одночасно вноситься і по батьківській, і з материнською сторін.
Перше дослідження такого роду було проведено з використанням мітохондріальної ДНК. Порівнюючи дані аборигенів всіх континентів, вчені виявили, що різноманітність мтДНК вище за все в Південній Африці. Більше того, там були виявлені такі типи, які ніде більше не зустрічалися. Вони оцінювалися за нуклеотидної складом як найбільш древні. Мітохондріальне ДНК населення інших континентів були менш різноманітні, їх порівняння з мтДНК аборигенів Південної Африки показало, що вони виникли як мутаційні зміни африканських типів у ході розповсюдження людства за межі Африки.
Вивчення географічного розподілу типів ДНК-маркерів і аналіз їх мутаційних взаємозв'язків дозволили довести справедливість монофілетичної гіпотези (рис. 4). Більш того, їх вивчення допомагає реконструювати міграційні події не тільки далеких тисячоліть, а й останніх століть. Наприклад, епоха великих географічних відкриттів ХIV-XVI ст. сприяла розвитку контактів з невідомими раніше мешканцями віддалених земель. На кораблях були тільки чоловіки, і зараз генетичні дослідження аборигенних народів Африки, Океанії і Америки показуючиють присутність в їх ДНК значної частки типів Y-хромосоми, характерних для європейців.
«Мітохондріальна Єва» і «Y-хромосомний Адам»
За ступенем різноманітності ДНК-маркерів можна встановити, від якого предкової типу і коли вони відбулися. Більше того, оскільки різні типи мтДНК НЕ рекомбінують, тобто не обмінюються між собою фрагментами в процесі утворення статевих клітин, то імовірнісний аналіз показує, що всі нині існуючі типи мтДНК зводяться в далекому минулому до одного-єдиного прототипу. Чому це відбувається? Припустимо, існувала невелика прапопуляція. Якщо одні жінки в ній мали більше дітей, ніж інші, то в наступному поколінні саме їх мтДНК буде зустрічатися частіше. Їх дочки і внучки теж мали дітей, унаследовавшх мтДНК своїх предків. При цьому випадковим чином виникають мутації. Так розвивається стохастичний процес у передачі типів мтДНК, в результаті чого з покоління в покоління одні типи мтДНК збільшують свою показність в популяції, інші - зменшують.
Згідно теорії стохастичних процесів, коли-небудь в популяції залишаться нащадки тільки однієї мтДНК - від якоїсь єдиної жінки, образно зветься "Мітохондріальної Євою". Але і її одноплемінника внесли свій внесок у генофонд віддалених нащадків, оскільки наш геном містить не тільки мітохондріальну ДНК. Довжина мтДНК лише близько 16,5 тис. нуклеотидів, що мізерно мало в порівнянні з 3 млрд. нуклеотидів ДНК хромосом, основна частина яких представлена 22 аутосомами і X-хромосомою, де комбінується спадковість по обох лініях, жіночого та чоловічого. Через численні мутацій мтДНК нині живих людей відрізняється від даних "Єви", вік якої (тобто точки з'єднання всіх типів мтДНК в минулому) приблизно 200 тисяч років. Аналогічна ситуація і з Y-хромосомою. Більша частина її НЕ рекомбінує, і мінливість ДНК в ній підкоряється тим же стохастичним процесам, що і мтДНК. При цьому говорять про "Y-хромосомному Адамі". Він виявився набагато молодший "мтЕви". Пояснення тому - менша, т. зв. ефективна кількість чоловіків протягом історії людства через їх більшої смертності і нерідко відсторонення від процесу розмноження.
неандерталський людина - наш прародитель або двоюрідний дядько? Додаткові відомості про нашу родоводу були отримані при порівнянні мітохондріальних ДНК сучасного людини, і неандертальця. Більше ста років, з тих пір, як в Німеччині вперше були знайдені останки нашого стародавнього родича, йшли дискусії про те, ким він нам доводиться. Виходячи з особливостей побудови кістяка неандертальця і його географічної поширеності, одні вчені вважали його нашим предком, тобто лінією Homo sapiens, розвиток якої призвело до людини сучасного анатомічного типу. Інші деталі дозволяли вважати його незалежної еволюційної гілкою, підвидом Homo sapiens, що має спільного з нами предка, тобто як би двоюрідним дядьком. Два підвиди Ці отримали зоологічне назву Homo sapiens neanderthalensis і Homo sapiens sapiens.
Ученим вдалося прочитати частину варіабельний (контрольного) ділянки мітохондріальної ДНК двох неандертальців. Перший був знайдений в Фельдховеровской печері в Німеччині, трохи пізніше був прочитаний генетичний текст мтДНК неандертальця-дитини, виявленого на Північному Кавказі в Мезмайской печері. При порівнянні найбільш мінливих фрагментів мтДНК сучасної людини, і неандертальця були знайдені істотні відмінності: вони відрізнялися один від одного в середньому по 27 нуклеотидних позиціями із 370 досліджених. Якщо порівняти мітохондріальної ДНК двох сучасних людей, то середня різниця буде дорівнює лише 8 нуклеотиду. Ці розрахунки показують, що наш спільний з неандертальцем предок жив приблизно 500-700 тис. років тому.
Аналіз ДНК дозволив зробити висновок, що обмін генами між людиною і неандертальцем не відбувався або був дуже малий. Найімовірніше, це зовсім окремі, паралельні еволюційні гілки, що відбулися від загального предка. Хоча для остаточного виведення потрібні додаткові дослідження ДНК. Так що неандерталець нам, схоже, еволюційний "дядя". Приблизно 300-400 тис. років тому відбулася остаточне розділення двох гілок. Неандертальці першими розселилися по Європі та Азії, потім туди прийшли люди сучасного типу (т.з. кроманьонскій чоловік), і вони досить довго співіснували на одній території. Але близько 30 тис. років тому неандерталський людина зникла, жодних його слідів в більш пізніх археологічних шарах знайдено не було. Можливо, він не витримав конкуренції і був витіснений і знищений своїм більш розумним і хитрим родичем, а може бути, існували й інші причини загибелі неандертальців.
Еволюція популяцій і рас
Вивчаючи різноманітність ДНК сучасних народів, можна оцінити чисельність тієї прапопуляціі, від якої, відповідно до гіпотези африканського походження, відбулося все людство. Вона була невелика - порядку декількох тисяч. Зіставляючи ДНК-маркери аборигенів Південної Африки, можна сказати, що приблизно 70-150 тис. років тому почалася інтенсивна диференціація і складні демографічні процеси, що супроводжуються виникненням різноманітних популяцій в межах Африки. Потім, 50-100 тис. років тому, хвилі переселенців стали вихлюпується за межі Африки і розтікатися по інших континентах, що відбилося на своєрідній структурі ДНК-древа (рис. 6).
Досліджуючи сучасне населення Європи, Азії, Океанії, Північної і Південної Америки і знаючи особливості та швидкість мутірованія в досліджуваних ДНК-маркери, можна з певною ступенем точності простежити шляхи і час міграцій людей з Африки. Дивно те, що генетичні дані підтверджуються археологічними знахідками. Наприклад, структура ДНК свідчить про те, що людина з'явилася в Австралії і Новій Гвінеї 50-60 тис. років тому. Аналіз складу хімічних елементів артефактів вказує на той же період. До Центральної і Південно-Східну Азію люди прийшли приблизно 70 тис. років тому, заселення Європи відбулося пізніше, 35-40 тис. років тому. Час освоєння Америки до цих пір не визначено, відомо лише, що люди з'явилися там набагато пізніше, ніж на інших континентах, - від 15 до 35 тис. років тому.
Як виникли сучасні раси людини і чи відрізняються вони один від одного по ДНК? Протягом десятків тисяч років йшли процеси міграцій та адаптації людини до місцевих умов. Припустімо, група людей прийшла у Південно-Східну Азію й осіла там на багато поколінь. Потім частина емігрувала далі, утворюючи нову локальну популяцію, яка, однак, має спільну історію і загальних предків з батьківського групою, а тому їх ДНК більш подібні між собою, ніж з жителями інших континентів. Дійсно, населення різних материків еволюційно набагато далі від загальної предкової групи, ніж сусідні популяції, близькі за родинним зв'язкам і демографічної історії. За той час, що минуло з часу відокремлення від загальних предків, їх ДНК стали відрізнятися один від одного за рахунок накопичуються у низці поколінь мутацій. Генетичні відмінності між людьми з різних материків ми називаємо сьогодні расовими ознаками. Вивчаючи десятки і сотні ДНК-маркерів, можна майже стовідсотково ідентифікувати расу (мал. 7). Щоб достовірно визначити етнічну приналежність індивіда в межах раси і великого географічного регіону, будуть потрібні тисячі ДНК-маркерів. А в зонах контакту різних рас і етнічних груп це зробити практично неможливо через змішання генофондів.
Але генетично ми всі вийшли з одного гнізда, причому порівняно недавно в масштабах еволюції (рис. 6).
Подальше розвиток рас йшло незалежно один від одного: люди адаптувалися до клімато-географічних умовами, типу харчування і ландшафту, складалися мова і культура. Але на формування народів впливали не тільки процеси розподілу популяцій. Нові етноси могли утворитися при змішуванні груп різної расової та мовної приналежності. При цьому виникала генетично різнорідна етнічна спільність з єдиним типом культури і спільною мовою. Тому зараз все більшої актуальності набувають дослідження, пов'язані з вивченням генофонду, тобто всього розмаїття ДНК в популяціях, генетичної історії населення окремих регіонів, расово-етнічних груп, родоводу сучасних етносів.
Контакт етносів
Особливий інтерес з цієї точки зору являє Волго-Уральський регіон - у силу особливостей етнічної історії населяють її народів. Тут зустрілися дві хвилі розселення, дві раси: європеоїдна і монголоїдна. Сліди цієї події зберігає ДНК проживаючих тут народів (рис. 8). Дослідження мітохондріальної ДНК та Y-хромосоми дозволили розрахувати час формування народів, що заселили згодом цей регіон. Це сталося приблизно 40-50 тис. років тому, що відповідає часу появи сучасної людини на європейському континенті в епоху верхнього палеоліту.
Порівняльний аналіз мтДНК 18 народностей Євразії, що відносяться до тюркської гілки алтайської мовної сім'ї, дозволив встановити західно-східний градієнт збільшення частоти азіатських типів мтДНК на просторі 8 тис. км: від 1% у гагаузів з Молдови до 95-99% у якутів і долган. Відповідно, європеоїдну риси найбільш властиві жителям Західної Євразії, а найменше - населенню Східного Сибіру. Народи Волго-Уральського регіону, а також узбеки і казахи, тобто ті, хто живе на кордоні між Європою та Азією, займають проміжне положення. Вивчення аутосомним ДНК-маркерів показало наявність в генофонд народів Волго-Уральського регіону значної частки європеоїдну рис - від 50 до 90%. Таким чином, опинившись на кордоні між Європою та Азією, ці народи зберегли сліди змішання двох рас, що прийшли один - зі Сходу, а інша - із Заходу. Крім того, виявилося, що подібність мов відіграє меншу роль, ніж географічна близькість популяцій. Якщо, наприклад, росіяни з Рязанської і Курської областей мають тільки 2-3% монголоїдні типів мтДНК, то росіяни, що проживають на кордоні Європи та Азії, мають їх уже 10-12%. Це пояснюється їх змішуванням з тюркомовними народами на території Волго-Уральського регіону.
Таким чином, ДНК якого-небудь індивіда не дає можливості визначити його національність, але дозволяє з'ясувати, якого типу в нього мтДНК або Y-хромосома: скажімо, монголоїдної або європеоїдної лінії. У росіян Волго-Уральського регіону є ДНК-маркери, які характерні і для марійців, і для мордви, і для чувашів, і для башкирів, і для татар, і для удмуртів; відповідно, марійці мають ДНК-маркери, що зустрічаються в інших етнічних груп Волго-Уральського регіону, і т. д. Ці результати показують глибоку генетичну спільність давно живуть поруч народів, хоча кажуть вони на несхожих мовами, вірять в різних богів і відрізняються культурними традиціями.
Всі люди - генетичні кузени і кузини
Ми все генетично надзвичайно схожі. Нас відрізняє один від одного ряд ознак, яким ми схильні надавати занадто велике значення (ріст, колір шкіри, форма голови та ін), але як вони нікчемні в порівнянні з нашим майже стовідсотковим (99,9%) генетичним схожістю! Ми сформувалися під впливом не тільки генів, але й оточуючих нас людей і явищ, причому у становленні особистості середу грає незмірно більшу роль, ніж спадкові особливості. Ми всі - одна велика генетична сім'я, яка живе на спільній планеті. І всі чвари між людьми виникають на побутовому грунті: з-за недотримання елементарних норм спільного проживання народів, неповаги до цінностей, особливостям і позірним дивацтв один одного.
Але уявіть себе на безлюдному острові, куди випадок закинув ще одного людини - зовсім іншої раси, іншого віросповідання, зі своєю мовою і звичками, - 99,9% генетичної схожості вас тут же об'єднають.
Розділ 4. Позатропічна прабатьківщина людства
Гіпóтеза позатропíчного похóдження люд́ини (або Гіпотеза позатропічної прабатьківщини людини) стверджує, що істинною прабатьківщиною Людини умілої (лат. Homo habilis) була не Африка, а інший, холодніший і суворіший регіон Землі.
Вперше гіпотезу про походження людини в умовах холодного клімату високих широт Євразії висунув в 1870-х роках німецький мандрівник і натураліст Моріц Веґенер, сучасник Чарльза Дарвіна. Він вважав, що саме суворі, несприятливі умови могли «примусити» деяких найрозумніших і діяльних мавп не загинути, а «олюднитися».
Протягом довгого часу гіпотеза залишалася в забутті, перш за все, через відсутність будь-яких речових доказів. Однак, нещодавно вона реанімувалася сенсаційним відкриттям на території Якутії стоянок часів раннього палеоліту — дирінгської культури. За первинними оцінками, вік цієї культури налічував 1,8-3,2 мільйони років, що мало відрізняється від віку знахідок в Олдувайській ущелині у Східній Африці.
Згідно з останніми дослідженнями, вік знарядь дирінгской культури виявився все ж таки значно менше перших оцінок і склав 260-370 тисяч років. Таким чином, гіпотеза позатропічної прабатьківщини людства знов виявилася позбавленою прямих археологічних доказів.
З моменту, як європейці проникли до Нового Світу, допитливі стали міркувати, звідки відбувається корінне населення континенту. Екзотичний вигляд медноліцих, прикрашених пір'ям аборигенів породжував чимало припущень. У 1787 р. майбутній президент США і (рідкісне поєднання) один з перших американських археологів Томас Джефферсон висловив думку про подібність індіанців з мешканцями Східної Азії і про ймовірність їх азіатських витоків. Тієї ж проблемою зацікавилися і російські мандрівники, освоюючи територію Нового Світу з боку Тихого океану. Один з учасників експедицій Берінга, натураліст Георг Стеллер, вважав, що колись Чукотка і Аляска з'єднувалися сушею між собою і мешканці Камчатки і Америки "походять від одного покоління".
Все це, зрозуміло, довгий час залишалося умоглядними припущеннями. Під враженням знахідок викопної людини в Європі американські вчені і любителі археології в кінці XIX ст. з ентузіазмом зайнялися пошуками власного палеоліту. На жаль, кожного разу виявлялося, що "палеолітичні давнини" відносяться до значно пізніших періодів. Серія розчарувань породила скептичне ставлення до самої можливості знайти сліди людини льодовикового періоду в Новому Світі. Лише в 20-ті роки, коли вдалося виявити кам'яні наконечники разом з кістками бізона вимерлого виду, реальність американського палеоліту стала очевидною навіть запеклим скептикам. Для позначення ранніх культур був запропонований термін "палеоіндейци".
З тих пір зусиллями кількох поколінь археологів в Новому Світі відкриті сотні стоянок прадавньої людини. Ботанік Ерік Хюльтен, який вивчав подібні види рослинного світу по обидві сторони Тихого океану, ввів у науку поняття "Берингія", що означає існувала колись сушу, що зв'язувала в єдине ціле Азію й Америку. У свою чергу вітчизняні дослідники первісності чимало потрудилися над вивченням палеоліту в різних куточках Сибіру і Далекого Сходу. Північноамериканські та російські археологи випробовують давній взаємний інтерес один до одного. Свідчення тому - численні публікації, ряд успішно проведених конференцій і спільних експедицій.
Коли і якими шляхами чоловік проник на територію Нового Світу? Чи можна знайти в Азії предків американських індіанців?
Це складна комплексна завдання, для вирішення якої необхідно об'єднати зусилля представників різних наук. Перш за все нам не обійтися без відомостей з четвертинної геології та палеогеографії, створюють ту естественноисторическую основу, без якої неможлива реальна реконструкція ранньої історії людства. Вигляд нашої планети 10-12 тис. років тому разючим чином відрізнявся від сучасного. Величезні простори півночі Євразії та Америки були покриті льодовиковими щитами, що вбирає в себе масу води. За рахунок цього рівень світового океану падав, і великі простори сучасних морів і проток осушувалися, створюючи свого роду сухопутні мости між континентами.
Здавалося б, для розкриття таємниць походження індіанців достатньо звернутися до кісткових останків самих древніх людей. На жаль, антропологічні знахідки льодовикового періоду одиничні і фрагментарні як у Сибіру, так і в Америці. Цікаві результати отримані при порівнянні генетики сучасних і древніх популяцій людини в Азії та Америці. * Про це докладно писали М. В. Деренко і Б. А. Малярчук у збірнику "Російська наука: грані творчості намежі століть". М., 2000. С.428-433.
Генетичні дані свідчать на користь походження індіанців від однієї предкової групи, найбільш близькою до сучасних монголоїдів. Однак оцінки часу існування такої групи вельми суперечливі. Деяке світло на питання початкового заселення Нового Світу можуть пролити результати зіставлення мов та міфології корінних народів Азії та Америки. Але тут знову нас чекає розчарування - адже мова йде про непрямі свідчення, не прив'язаних до хронології і не піддаються однозначній інтерпретації. Досить сказати, що лінгвісти до цих пір не прийшли до єдиної думки про те, скільки ж мовних сімей налічується у індійців.
Залишаються єдино достовірні, хоча і "німі", сліди минулого - археологічні пам'ятники, залишки стійбищ далеких предків. До них ми і звернемося.