

1.4. Участие спинного мозга в регуляции тонуса мышц
Регуляция тонуса мышц имеет рефлекторную природу и осуществляется посредством миотатических и позно-тонических рефлексов.
Миотатический рефлекс является рефлексом растяжения, который начинается с раздражения мышечного веретена. Практически в каждой мышце находятся мышечные рецепторы растяжения, называемые из-за своей формы «мышечными веретенами» (рис.2).
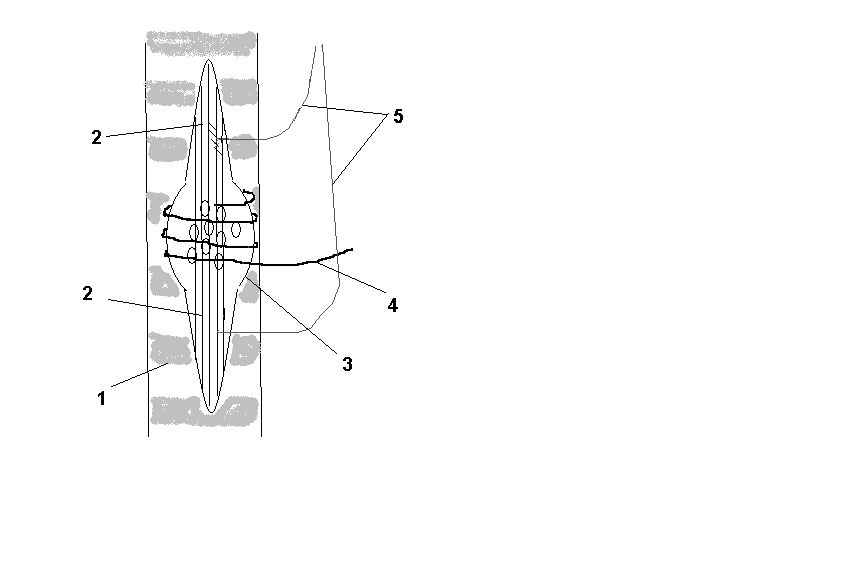
Рис.2. Схема мышечного веретена.
1 – экстрафузальные волокна; 2 – интрафузальные волокна; 3 – ядерная сумка;
4 – чувствительные нервные волокна; 5 – эфферентные волокна гамма-мотонейронов
Мышечный рецептор состоит из поперечнополосатых интрафузальных мышечных волокон, окруженных соединительнотканной капсулой, и имеет форму веретена. Длина интрафузальных мышечных волокон равна 4 - 7 мм, толщина 15 — 30 мкм. Мышечные веретена располагаются параллельно скелетной мышце и своими концами крепятся к соединительнотканной оболочке пучка экстрафузальных мышечных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5 - 1 мм. Имеется два типа мышечных веретен, различающихся по расположению ядер. В одних веретенах ядра сконцентрированы в центре и называются волокнами с ядерной сумкой. В других — ядра в средней части волокна располагаются цепочкой и называются волокнами с ядерной цепочкой. Вокруг средней части мышечного веретена несколько раз обвивается окончание афферентного нервного волокна группы Iа. Некоторые мышечные веретена обвиваются афферентным волокном группы II на периферическом участке (преимущественно в волокнах с ядерной цепочкой).
Механизм регуляции тонуса мышц представлен на рисунке 3. Растяжение экстрафузальных мышечных волокон (при расслаблении мышцы) приводит к растяжению мышечного веретена. При растяжении мышечного веретена происходит растяжение ядерной сумки и возбуждение рецептора растяжения. Импульсы поступают в спинной мозг к α-мотонейронам. Возбуждение по аксонам α-мотонейронов вызывает сокращение экстрафузальнох мышечных волокон.
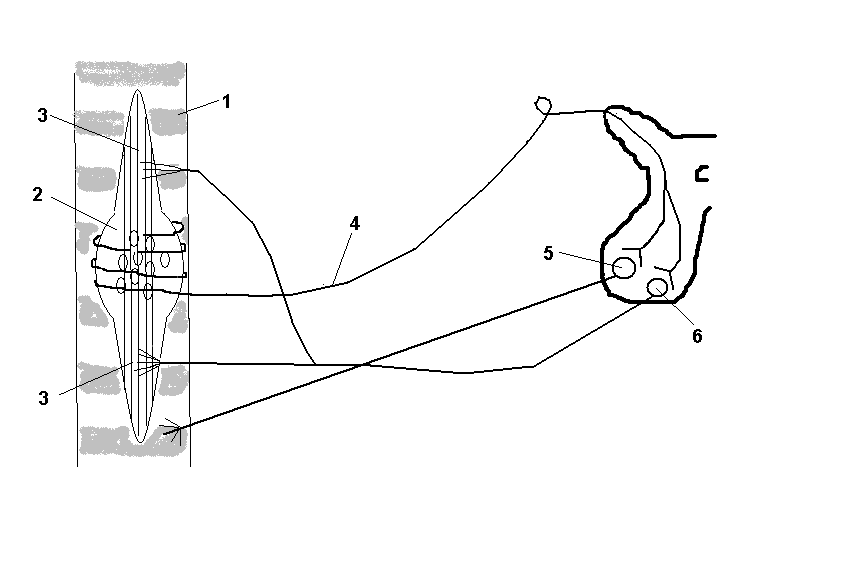
Рис.3. Схема регуляции мышечного тонуса.
1 – экстрафузальные волокна; 2 – мышечное веретено; 3 – интрафузальные
волокна; 4 – эфферентные нервные волокна; 5 – альфа-мотонейроны;
6 – гамма-мотонейроны
Гамма-петля: Ряд воздействий от рецепторов мышц и сухожилий идет не только на α-мотонейроны, но и по коллатерали аксона на γ-мотонейроны: γ-мотонейроны возбуждаются → посылают импульсы к интрафузальным волокнам → интрафузальные волокна сокращаются → мышечное веретено растягивается → рецептор возбуждается → импульсы идут в спинной мозг → активация α-мотонейронов → сокращение экстрафузальных волокон.
Позно-тонические рефлексы спинного мозга направлены на поддержание позы. С их помощью регулируется тонус мышц. Эти рефлексы возникают с проприорецепторов мышц и фасций шеи. Переключение с этих рецепторов осуществляется на уровне шейного отдела спинного мозга и приводит к изменению тонуса мышц при изменении положения головы и шеи.
1.5. Сегментарный и межсегментарный принцип работы спинного мозга
Спинной мозг состоит из 31-32 сегментов. Каждому сегменту соответствуют две пары корешков (соответственно числу сегментов из спинного мозга выходит 31-32 пара передних двигательных корешков и входит в него 31-32 пары задних чувствительных корешков). При проекции сегментов спинного мозга на позвонки приходится учитывать несоответствие длины спинного мозга и позвоночника. В шейном отделе сегменты расположены на 1 позвонок выше, чем соответствующий им по счету позвонок; верхнегрудные – на 2, нижнегрудные – на 3 (пример: V шейный сегмент расположен на уровне IV шейного позвонка, V грудной – на уровне III грудного позвонка, XI грудной – на уровне VIII грудного позвонка и т.д.). В большом несоответствии находятся поясничные и крестцовые сегменты и позвонки: поясничные сегменты находятся на уровне X, XI, и XII грудных позвонков; крестцовые – XII грудного и I поясничного. Морфологических границ между сегментами спинного мозга нет, поэтому деление на сегменты является функциональным и определяется зоной распределения в сегменте волокон заднего корешка и зоной клеток, которые образуют передний корешок. Каждый сегмент иннервирует через свои корешки 3 метамера тела и получает информацию от 3 метамеров тела. В итоге перекрытия нервных волокон каждый метамер тела иннервируется тремя сегментами и передает сигналы в 3 сегмента спинного мозга, что повышает надежность регуляторных механизмов.