

Механизм действия нестероидных гормонов
М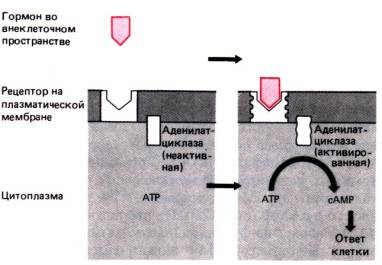 еханизм
действия гормона, образующего комплекс
с рецептором, локализованным на
плазматической мембране. Достигнув
рецептора и взаимодействуя с ним, гормон
вызывает его трансформацию. Трансформация
приводит к активации аденилатциклазы,
также локализованной в плазматической
мембране. Аденилатциклаза катализирует
дефосфорилирование АТР с образованием
сАМР. сАМР вызывает в клетке разнообразные
эффекты, которые приводят в конечном
счете к физиологическому ответу клетки.
(АТР–аденозинтрифосфат, АТФ;
сАМР–циклический аденозинмонофосфат,
цАМФ)
еханизм
действия гормона, образующего комплекс
с рецептором, локализованным на
плазматической мембране. Достигнув
рецептора и взаимодействуя с ним, гормон
вызывает его трансформацию. Трансформация
приводит к активации аденилатциклазы,
также локализованной в плазматической
мембране. Аденилатциклаза катализирует
дефосфорилирование АТР с образованием
сАМР. сАМР вызывает в клетке разнообразные
эффекты, которые приводят в конечном
счете к физиологическому ответу клетки.
(АТР–аденозинтрифосфат, АТФ;
сАМР–циклический аденозинмонофосфат,
цАМФ)
Рецептор локализован на поверхности мембраны и реакция клеток-эфекторов реализуется через циклазную систему, т.е. циклическая 3,5 АМФ формируется в 1 АТФ под влянием адениилатциклазы, а инактивируется фосфодиэстилазой.
Г. взаимодействует с рецептором, сигнал об этом взаимодействии передается на адениилатциклазу, локализованную на внутренней поверхности мембраны, или на фосфодиэстилазу.
При этом увеличивается или уменьшается образование циклического 3,5 АМФ, ктр в свою очередь активирует внутриклеточные ферменты.
Таким образом, именно 3,5 АМФ является тем внутриклеточным посредником, ктр обеспечивает передачу влияния г. на внутриклеточные системы. Характер же конечного ответа клетки определяется спецификой ферментного набора эффекторной клетки, а действие г. фактически оказывается толчком пускающим в ход реакцию предопределенную ферментативным профилем клетки.
Белковые гормоны, факторы роста, нейротрансмиттеры, катехоламины и простагландины относятся к группе гормонов, для которых рецепторы расположены на плазматических мембранах клеток.
Механизм действия стероидных гормонов
В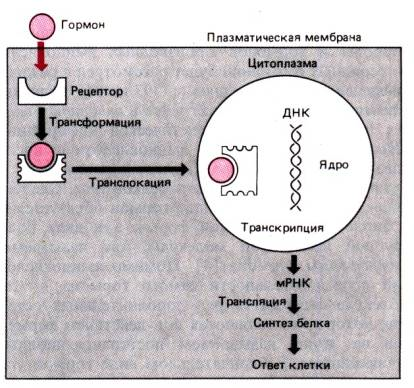 заимодействие
гормона с внутриклеточным рецептором.
Гормон диффундирует через плазматическую
мембрану и взаимодействует с рецептором.
Гормон–рецепторный комплекс переносится
в ядро и действует на синтез ДНК, изменяя
скорость транскрипции и количество
информационной (матричной) РНК (мРНК).
Увеличение или уменьшение количества
мРНК влияет на синтез белка в процессе
трансляции, что приводит к изменению
функциональной активности клетки
заимодействие
гормона с внутриклеточным рецептором.
Гормон диффундирует через плазматическую
мембрану и взаимодействует с рецептором.
Гормон–рецепторный комплекс переносится
в ядро и действует на синтез ДНК, изменяя
скорость транскрипции и количество
информационной (матричной) РНК (мРНК).
Увеличение или уменьшение количества
мРНК влияет на синтез белка в процессе
трансляции, что приводит к изменению
функциональной активности клетки
Циркулируя в крови стероиды присутствуют в виде свободных связанных форм. Белками плазмы связаны 98% половых гормонов, 90% кортизола, 50% альдостерона.
Гормон, находящийся в свободной форме, способен взаимодействовать с мембраной клетки и проходить через нее в цитоплазму, где связывается с цитоплазматическим рецептором, который отличается высокой специфичностью. Например, из гепатоцитов выделены рецепторные белки, связывающие только глюкокортикоидные гормоны или эстрогены. В настоящее время идентифицированы рецепторы к эстрадиолу, андрогенам, прогестерону, глюкокортикоидам, минералокортикоидам, витамину Д, тироидным гормонам, а также к ретиноивой кислоте и некоторым другим соединениям.
Поступление стероидов в клетку не зависит от мембранных барьеров или процессов транспорта.
Стероид в начале связывается в цитоплазме с плазматическим рецептором, ктр подвергается активации и комплекс рецептор-гормон перемещается из цитоплазмы в ядро и уже здесь происходит связывание комплекса ядерными акцепторными участками хроматина клеток-эфекторов.Это модулирует активность клеток специфических генов, ответственных за синтез определенных матричных РНК.
Механизм действия стероидных (жирорастворимых) гормонов
Проникновение стероида (С) в клетку
Образование комплекса СР (стероид-рецептор). Все Р стероидных гормонов представляют собой глобулярные белки примерно одинакового размера, с очень высоким сродством связывающие гормоны
Трансформация СР в форму, способную связываться ядерными акцепторами [СР] Любая клетка содержит всю генетическую информацию. Однако при специализации клетки большая часть ДНК лишается возможности быть матрицей для синтеза иРНК. Это достигается путем сворачивания вокруг белков гистонов, что ведет к препятствию транскрипции. В связи с этим генетический материал клетки можно разделить на ДНК 3-х видов: 1.транскрипционно неактивная 2.постоянно экспрессируемая 3.индуцируемая гормонами или другими сигнальными молекулами.
Связывание [СР] с хроматиновым акцептором. Следует отметить, что этот этап действия С полностью не изучен и имеет ряд спорных моментов. Считается что [СР] взаимодействует со специфическими участками ДНК так, что это дает возможность РНК-полимеразе вступить в контакт к определенным доменам ДНК.
Интересным является опыт, который показал, что период полужизни иРНК при стимуляции гормоном увеличивается. Это приводит к многим противоречиям: становится непонятно увеличение количества иРНК свидетельствует, о том что [СР] повышает скорость транскрипции или увеличивает период полужизни иРНК; в то же время увеличение полужизни иРНК объясняется наличием большого числа рибосом в гормон-стимулированной клетке, которые стабилизируют иРНК или другим действием [СР] неизвестным для нас на сегодняшний момент.
Избирательная инициация транскрипции специфических иРНК; координированный синтез тРНК и рРНК Можно полагать, что основной эффект [СР] состоит в разрыхлении конденсированного хроматина, что ведет к открыванию доступа к нему молекул РНК-полимеразы. Повышение количества иРНК приводит к увеличению синтеза тРНК и рРНК.
Процессинг первичных РНК
Транспорт мРНК в цитоплазму
Синтез белка
Посттрансляционная модификация белка
Стероиды вначале связываются специфическими белками мембраны клетки, которые транспортируют их к цитоплазматическому рецептору или, минуя его, непосредственно к рецепторам ядра. Цитоплазматический рецептор состоит из двух субъединиц. В ядре клетки субъединица А, взаимодействуя с ДНК, триггирует (запускает) процесс транскрипции, а субъединица B связывается с негистоновыми белками. Эффект действия стероидных гормонов проявляется не сразу, а спустя определенное время, которое необходимо для образования РНК и последующего синтеза специфического белка.