

ЛАБОРАТОРНАЯ РАБОТА 1
ИССЛЕДОВАНИЕ ДЛИТЕЛЬНОГО ПОСЛЕСВЕЧЕНИЯ ЛИСТЬЕВ РАСТЕНИЙ
Цель: исследование влияния интенсивности света:
- различных участков видимого спектра (400-750 нм)
- температуры на интенсивность и кинематику послесвечения
Теоретическая часть:
Длительное послесвечение было открыто в 1951 году Арнольдом и Стреллером. Этот эффект состоит в том, что все фотосинтезирующие объекты (листья, водоросли, фотосинтезирующие бактерии) после предварительного освещения испускают свечение. Спектр испускания послесвечения у зеленых растений совпадает со спектром флуоресценции хлорофилла и это позволяет говорить, что излучение происходит при переходе молекул хлорофилла из синглетного возбужденного состояния в основное состояние S*→S0. Энергия, необходимая для возбуждения молекул хлорофилла, генерируется в ходе обратных реакций рекомбинации первичных продуктов фотосинтеза. Это свечение происходит с затуханием. Время жизни послесвечения составляет от 10-6 до 100 с и значительно превышает время жизни синглетного возбужденного состояния хлорофилла 10-10 – 10-9 с. В настоящее время весь процесс послесвечения зеленых растений разделяют на 5 компонент.
Первая компонента - время жизни 510-3 с, интерпретируется как замедленная флуоресценция хлорофилла " а ", связанная с функционированием II фотосистемы фотосинтеза..
Вторая компонента - время жизни 10-2 с, послесвечение возникает в ходе обратимых фотопроцессов, протекающих с участием пигментного комплекса, в который входит хлорофилл и каротин.
Третья компонента - время жизни 1,7 с, полагают, что фотопродукт, ответственный за послесвечение этого компонента образуется в пределах II фотосистемы.
Четвертая компонента - время жизни 1,59 с, полагают, что свечение связано с функционированием I фотосистемы.
Пятая компонента - время жизни 10 - 15 с и более возникает только при положительных температурах, интерпретируется как хемилюминесценция.
Поскольку процесс послесвечения включает несколько компонент, то это говорит об участии в процессе послесвечения нескольких фотопродуктов с различным временем жизни.
Таким образом, в основе быстрых компонент (1 - 4 ) послесвечения лежат элементарные фотофизические и фотохимические процессы, развертывающиеся в непосредственной близости от молекул пигмента, и излучение представляет собой обращение первичного запасания энергии световых квантов. Что касается долгоживущей компоненты ( 5 ) , то ее рассматривают как явление хемилюминесценции хлорофилла, которое связано с окислением относительно долгоживущих фотовосстановленных продуктов фотосинтеза.
Известно, что световая стадия фотосинтеза зеленых растений включает две пространственно разделенные фотохимические системы, сопряженные цепью окислительно-восстановительных переносчиков ( рис.1 ).
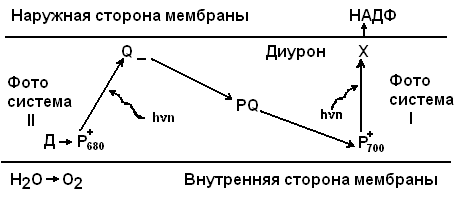
Рис.1. Схема электрон-транспортной цепи фотосинтеза растений в тилакоидной мембране.
Р680 и Р700 - молекулы пигмента ( хлорофилла ) реакционных центров фотосистем IIи I соответственно, Д- первичный донор, Q и X - первичные акцепторы, PQ - пул пластохинонов, диурон - ингибитор транспорта электронов.
Для зеленых растений показано, что послесвечение генерируется, в основном, в реакционном центре ( РЦ ) фотосистемы II ( ФС II ). При повреждении РЦ свечение не наблюдается.
Суть механизма генерации послесвечения у зеленых растений и его регуляции можно представить следующей гипотетической схемой ( Рис.2 }.
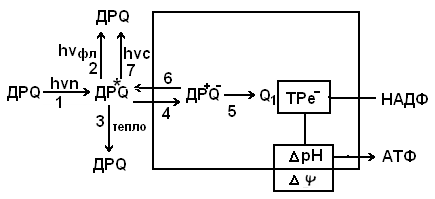
Рис.2. Схема генерации послесвечения и его регуляции.
ФС II - фотосистема II, Р - пигмент, Д и Q - первичные донор и акцептор ФС II, Тре- - транспорт электронов, ΔрН - протонный градиент, hvn, hvфл hvc - кванты поглощенного света, флуоресценции и послесвечения соответственно, 1 - 7 -переходы.
При поглощении кванта света (hvn) хлорофиллом Р реакционного центра фотосистемы II он переходит в возбужденное состояние Р* (переход 1). Из возбужденного состояния Р* может переходить либо в основное состояние Р с излучением кванта быстрой флуоресценции (переход 2), либо в основное состояние Р без излучения с выделением тепла (переход 3), либо происходит переход электрона на уровень первичного акцептора Q с образованием зарядов ДР+0", т.е. происходит первичное разделение зарядов в фотохимическом РЦ (переход 4). Далее может происходить захват электрона вторичным акцептором (переход 5).
Вместе с тем возможен переход электрона от первичного акцептора Q на Р ( рекомбинация зарядов ), т.е.обратный процесс с образованием возбужденного состояния Р+ (переход 6) с последующим испусканием кванта замедленной флуоресценции (переход 7).Таким образом, энергия для послесвечения у зеленых растений выделяется в реакциях рекомбинации окисленного пигмента Р+ реакционного центра и восстановленного акцептора Q-.
Однако считают, что послесвечение высвечивает не пигмент РЦ, а светособирающие молекулы хлорофилла реакционного центра.
Интенсивность послесвечения регулируется многими факторами и в первую очередь транспортом электронов. Установлено, что чем больше скорость транспорта электронов, тем выше интенсивность послесвечения. На этом принципе основаны рекомендации по использованию, послесвечения как теста для определения активности фотосистемы II в интактных клетках. Однако эта связь не всегда носит прямопропорциональный характер. В свою очередь эта связь зависит от работы фотосистемы I и восстановления НАДФ. Повышение активности фотосистемы I вызывает усиление оттока электронов от Q и приводит к снижению интенсивности послесвечения .Другой важной причиной, нарушающей пропорциональность между скоростью транспорта электронов и интенсивностью свечения является их зависимость от величины электрического потенциала ΔΨи протонного градиента ΔрН на мембране.
Поскольку послесвечение возникает в результате протекания первичных реакций фотосинтеза ( особенно короткоживущих компонент ) , то оно несет ценную информацию о функционировании фотосинтетического аппарата и о природе первичных продуктов фотосинтеза, образующихся в этом процессе. Поэтому метод регистрации послесвечения широко используют в научных исследованиях механизмов фотосинтеза.