

Лекция – 15
Редакция 11.10.2009 г.
Тема: «Коэффициент наследуемости»
Вопросы.
1. Дисперсия проявлений признаков в популяциях.
2. Коэффициент наследуемости в широком смысле.
3. Расчет величины коэффициента наследуемости в широком смысле при селекционной оценке популяций.
4. Вычисление значений коэффициента наследуемости в узком смысле.
Ключевые слова:
наследуемость, генотип, фенотип, коэффициент наследуемости, дисперсия, популяция, особь, признак, фенотипическая варианса, генотипическая варианса, средовая варианса, генетический анализ популяций.
Литература.
Вавилов Н.И.
Котов М.М. Генетика и селекция. Часть 1.: Учебник для вузов. – Йошкар-Ола: МарГТУ, 1997. – 280 с.
Котов М.М. Генетика и селекция. Часть 2.: Учебник для вузов. – Йошкар-Ола: МарГТУ, 1997. – 108 с.
Любавская А.Я. Лесная селекция и генетика.: Учебник для вузов. – М.: Лесная промышленность, 1982. – 288 с.
Бессчетнов В.П. Полиморфизм казахстанских популяций облепихи крушиновидной (Hippophae rhamnoides L.) по хозяйственным и адаптивным признакам. Дис. док. биол. наук. Алматы, 1994. – 543 с.
Бессчетнов В.П.
Чмыр А.Ф., Бессчетнов В.П. Экология и культура облепихи Hippophae rhamnoides L. – СПб, СПбНИИЛХ, 1998. – 278 с.
Джонатат В. Райт. Введение в лесную генетику. М.: Лесная промышленность, 1978. – 470 с.
Гужов Ю.Л., Фукс А., Валичек П. Селекция и семеноводство культурных растений. – М.: Агропромиздат, 1991. – 463 с. (Учебник для студентов высш. Учеб. Заведений).
1. Дисперсия проявлений признаков в популяциях Теоретическое обоснование
Условием результативности отбора является генотипическая обусловленность хотя бы части наблюдаемой фенотипической изменчивости признака, подлежащего отбору. Мы с вами останавливались на этих аспектах успешности отбора. Чем больше доля генотипически обусловленной изменчивости признака в его общей фенотипической изменчивости, тем сильнее связь между генотипом и фенотипом и, следовательно, тем эффективнее отбор. Абсолютная связь между генотипом и фенотипом в природных популяциях возможна только в гипотетическом случае, когда вся видимая фенотипическая изменчивость признака обусловлена генотипически. В этом случае достигается предельно высокая эффективность отбора (стопроцентная эффективность).
Добавить из Айала и др., 1988 Т.2 о соотношении генотипической и фенотипической долей с общей фенотипической дисперсии.
Мерой доли генотипически обусловленной изменчивости в общем фенотипическом варьировании служит коэффициент наследуемости.
Косвенно по коэффициенту наследуемости можно судить и о доле изменчивости, обусловленной влиянием среды: по разности между общей фенотипической изменчивостью и её долей, обусловленной генотипически.
П.Ф. Рокицкий (1978, стр. 201-202) считает, что предпосылкой для введения этого понятия в употребление при научном анализе строения популяций явился ряд обстоятельств. Оказалось, что применяемые на первоначальных этапах аналитической работы такие важнейшие биометрические показатели как «средняя величина признака в популяции» и «дисперсия проявлений признака в популяции», оцениваемая по среднему квадратическому отклонению, оказались недостаточными в работе с популяциями сложной структуры. В таких популяциях осуществляется большое число разнообразных скрещиваний, вследствие чего нет возможности оперировать анализом отдельных генов и пользоваться такими понятиями как «частота генов». И хотя с помощью оценок средних значений и дисперсии признаков удалось сделать существенные выводы об общей картине наследования количественных признаков и их генетической обусловленности отдельными генами, дальнейший анализ популяций требовал развития аналитических методов. Поэтому понадобились новые параметры для характеристики генетической структуры популяции по количественным признакам.
Подойти к пониманию принципа формирования новых методов анализа генетической структуры популяций можно следующим образом.
Любая варианта (значение признака) у какой-либо особи в популяции (например: значение диаметра ствола на высоте груди у конкретного дерева сосны и др.) имеет определенное фенотипическое значение «y» (например: диаметр равен y = 36 см). Это фенотипическое значение может быть выражено как отклонение от популяционной средней «М» (например: средний диаметр ствола в насаждении равен y =24 см) и может быть обозначено символом «Р». Тогда:
у – М = Р (например: 36 см – 24 см = +12 см, следовательно Р = 12 см).
Так как вариация значений признака в популяции определяется в конечном счете двумя основными факторами: различиями в генетической структуре отдельных особей и влиянием средовых факторов, – то и для каждой отдельной особи её фенотипическое значение Р можно рассматривать как результат суммирования генотипического (G) и средового (E) отклонений.
P = G + E
Тогда:
G = yG – MG
E = yE – ME
Но из этого, конечно, не следует, что в фенотипе каждой особи можно выделить самостоятельные доли, зависящие от генотипа яйца, из которого развился данный фенотип, и от условий среды, в которых он развился. Фенотип особи – это единое целостное понятие. О генотипе особи непосредственно, без анализа её потомства, судить невозможно. Выделение же долей вариации, зависящих от наследственности и от среды, производится только для популяций.
Чтобы определить вариансы (оценки дисперсии через среднее квадратическое отклонение) отклонений G и E для популяции, надо возвести каждое отклонение от среднего в квадрат, затем просуммировать все отклонения и полученные суммы разделить на число объектов в популяции (если учет был сплошным) или в выборке из популяции (при выборочных методах анализа). При этом необходимо учитывать установленное выше равенство P = G + E.
Дадим определения используемым понятиям.
Варианса – это мера дисперсии (изменчивости) признака у изучаемого объекта.
Дисперсия – это рассеивание значений признака, разброс значений признака.
Если мы возводим в квадрат левую половину равенства (именно для популяции), то должны возвести в квадрат и его правую половину для сохранения условия равенства, тогда получим:
P2 = (G + E)2 или P2 = G2 + E2 + 2GE
Поскольку речь идет о популяционном анализе, то суммируем значения отклонений всех особей. Получим несколько модифицированное выражение приведенного выше равенства.
ΣP2 = Σ(G + E)2 или ΣP2 = ΣG2 + ΣE2 + Σ2GE или ΣP2 = ΣG2 + ΣE2 + 2ΣGE
Далее, разделив каждый член равенства на численность объектов (в выборке или популяции) “n”, получим:
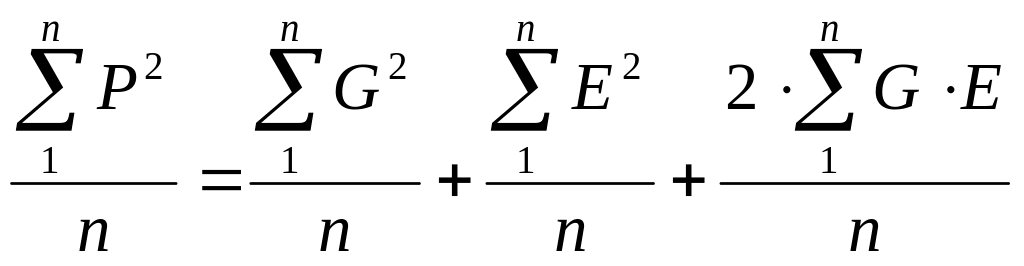
Но тогда мы сможем каждый из членов полученного равенства выразить в виде соответствующих дисперсий, а точнее, средних квадратов отклонений от среднего каждого конкретного значения признака, поскольку:
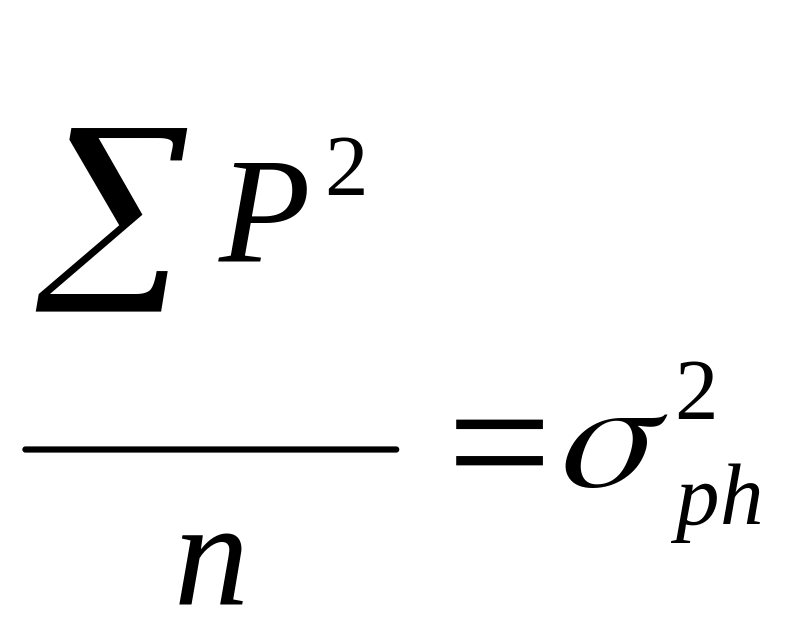 - общая фенотипическая
дисперсия;
- общая фенотипическая
дисперсия;
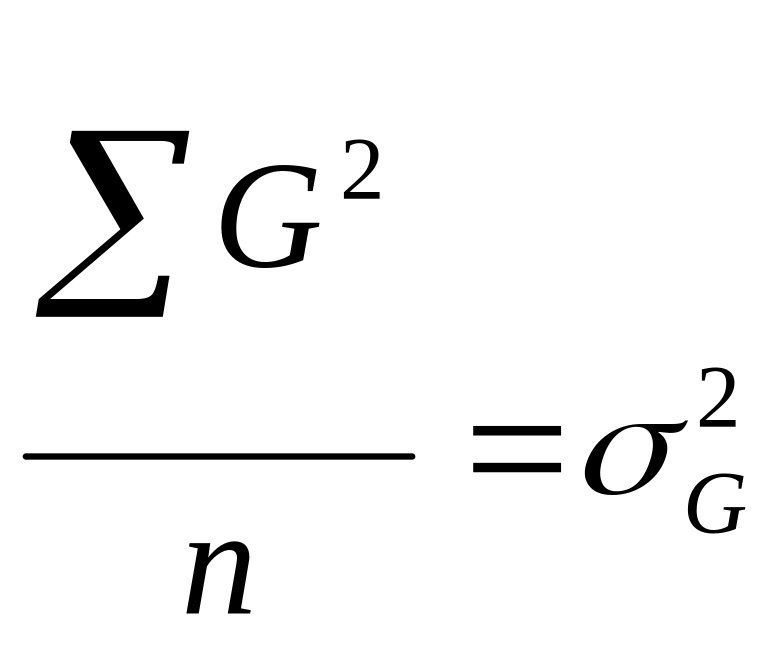 - генотипическая
дисперсия;
- генотипическая
дисперсия;
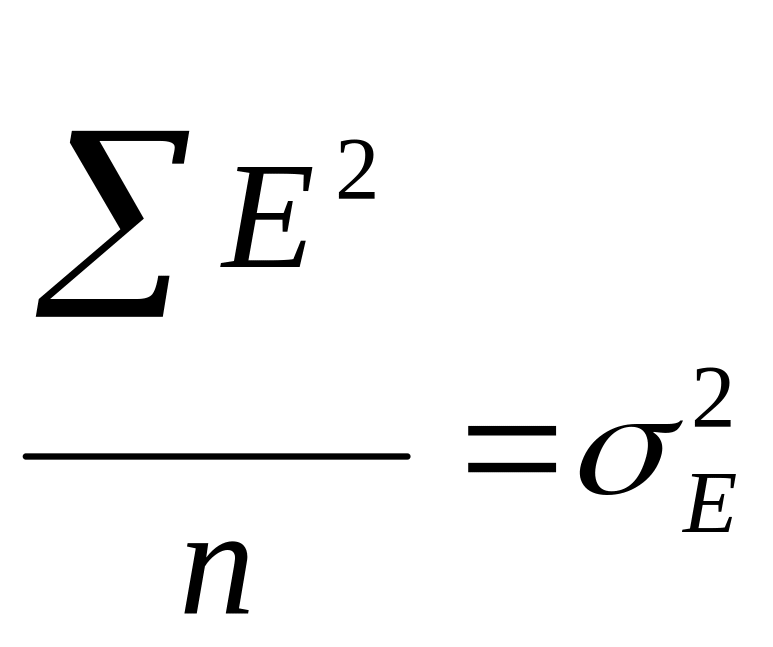 - дисперсия,
вызванная влиянием среды.
- дисперсия,
вызванная влиянием среды.
Сложнее стоит
вопрос о величине
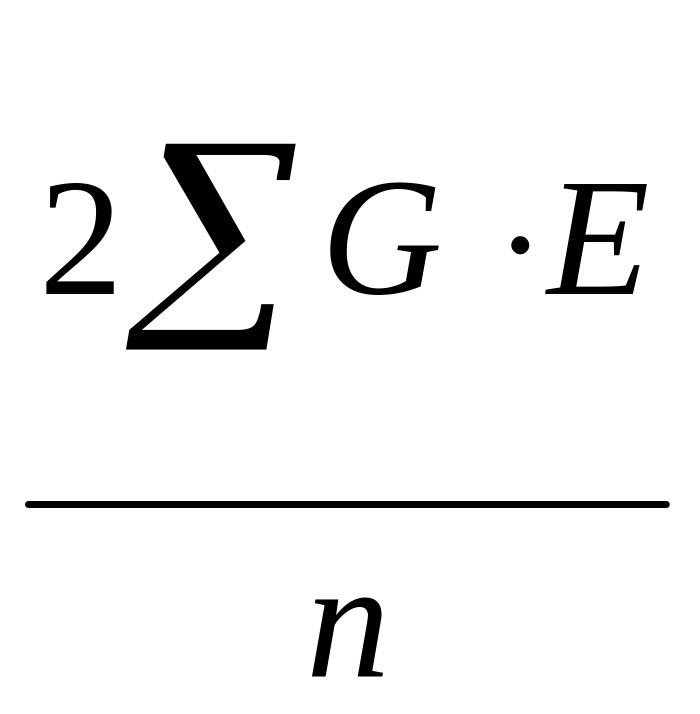 .
.
Ответить на него можно, исходя из следующих соображений. Поскольку величины G и E представляют собой генотипические и средовые отклонения, то сумма их произведений (сумма произведений отклонений значения от соответствующего среднего значения) является ковариансой между этими отклонениями.
![]()
Ранее мы приняли условие, согласно которому все величины в рассматриваемом равенстве P = G + E представляют собой отклонения от соответствующей средней величины оценок фенотипических проявлений признака в популяции, а именно:
у – М = Р
и соответствующих фенотипической оценке отклонений проявления признака, обусловленного генотипом
G = yG – MG
и вызванного действием факторов среды
E = yE – ME.
Как известно, коварианса входит в структуру коэффициента корреляции, а именно:
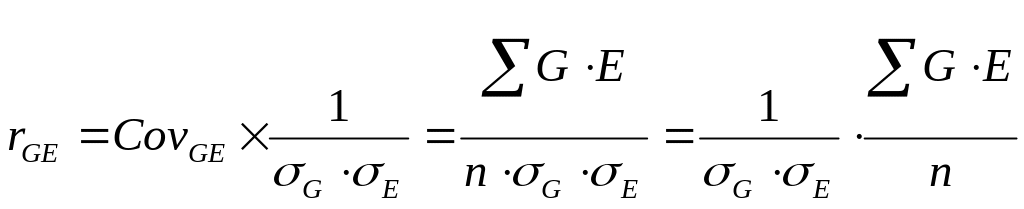
Тогда исходное выражение рассматриваемого компонента уравнения можно представить в следующем виде:

В таком случае первоначальное равенство
примет вид:
![]()
Изменения условий среды в пределах нормы реакции (и не связанные с эволюционными изменениями в чередующемся ряду поколений – этот аспект не рассматривается) не вызывают изменения генотипов особей вида, обитающих в границах территории, флуктуации экологических условий которой рассматриваются. Раметы одного клона, размещенные в разных местах на одной ЛСП, не изменят свой генотип в том или ином участке, как и в случае их размещения на других ЛСП. Тогда различия в генотипах разных клонов (клонов разных ПД) останутся стабильными и независимыми от условий среды.
Если предположить, что корреляция между вариацией генотипических и средовых факторов отсутствует или равна нулю, что вполне логично и вытекает из представлений о независимости в пределах нормы реакции варьирования, обусловленного генотипически, то формула примет знакомый нам вид:
![]() ,
где
,
где
![]() - общая фенотипическая
варианса;
- общая фенотипическая
варианса;
![]() - доля общей
фенотипической вариансы, зависящая от
различий между отдельными особями
популяции по их наследственности;
- доля общей
фенотипической вариансы, зависящая от
различий между отдельными особями
популяции по их наследственности;
![]() -
доля общей фенотипической вариансы,
вызванная различиями в условиях внешней
среды, при которых развивались особи
популяции.
-
доля общей фенотипической вариансы,
вызванная различиями в условиях внешней
среды, при которых развивались особи
популяции.
Соотношения между значениями генотипической и средовой варианс могут быть различными. Так если экспериментатор работает с клонами (черенковые саженцы, полученные от одного дерева), то в этом случае генотипическая варианса будет равна нулю, и общая фенотипическая варианса будет состоять только из средового компонента. Если же работа ведется на выровненном экофоне (в абсолютно контролируемых условиях фитотронов) с различным по происхождению материалом, то различия между сравниваемыми объектами будут определяться преимущественно (в идеале полностью) различиями в их генотипах.
Таким образом, получено наиболее общее выражение, характеризующее фенотипическую структуру популяции по количественному признаку. Фенотипическая варианса распадается на два компонента – генотипическую и средовую вариансы. P = G + E
Какими бы путями ни создавалась фенотипическая изменчивость в популяциях, и из каких бы компонентов она ни состояла, она всегда может быть выявлена и измерена вариансой – мерой оценки дисперсии. Самое существенное заключается в выяснении того, какую долю в общей фенотипической вариансе занимает варианса, зависящая от генетических различий между особями в популяции, то есть установлению величины отношения генотипически обусловленной вариансы к общей фенотипической вариансе. Такое отношение принято называть коэффициентом наследуемости.
Коэффициент наследуемости определяется соотношением варианс (дисперсий, средних квадратов отклонений), поскольку из обычных статистических мер рассеивания (дисперсии) только вариансу (2) можно разложить на компоненты.
С селекционно-генетической точки зрения фенотипическая варианса (оценка степени варьирования) в самом простом случае может быть обусловлена тремя причинами:
генотипическими различиями, обусловленными непосредственным действием генов (аддитивный компонент вариансы -
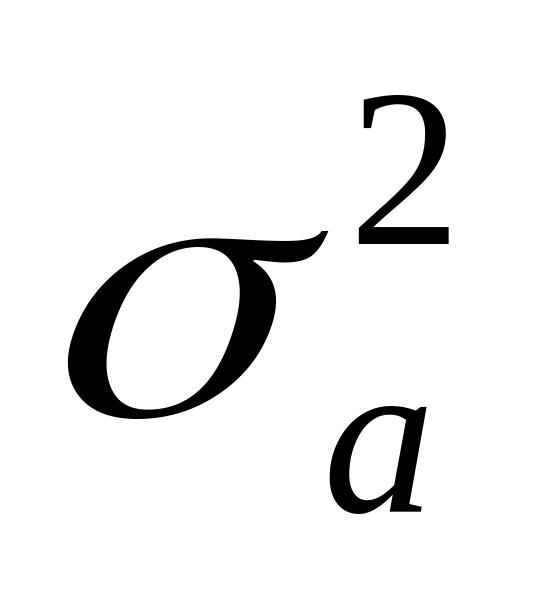 ,
не связанный с действием эффекта
доминирования);
,
не связанный с действием эффекта
доминирования);генотипическими различиями, возникающими вследствие межаллельных взаимодействий (компонент вариансы, обусловленный доминированием -
 );
);ненаследственными различиями между объектами, вызванными неодинаковым влиянием разнообразных локальных проявлений условий среды (компонент вариансы, обусловленный условиями внешней среды -
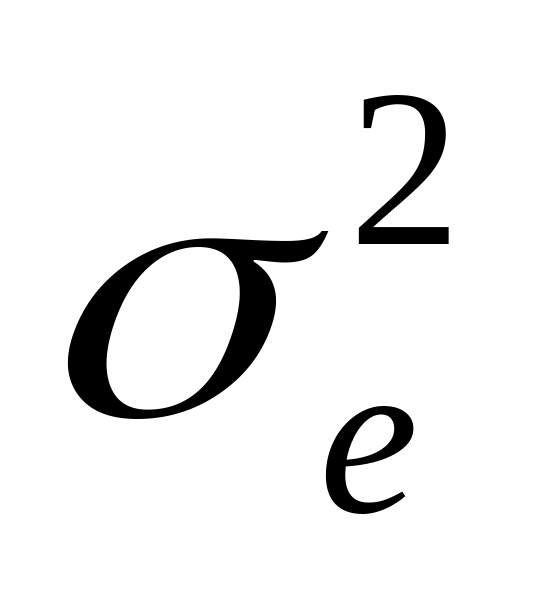 ).
).
Поскольку компоненты
вариансы суммируются, общая фенотипическая
варианса (![]() )
представляет их сумму. Общепринята
следующая формула её расчета:
)
представляет их сумму. Общепринята
следующая формула её расчета:
![]()
![]()
Так как аддитивная
генотипическая варианса и неаддитивная
генотипическая варианса являются
генотипическими компонентами
фенотипиеческой (общей) изменчивости,
то их можно суммировать и получить
генотипически обусловленную вариансу
(![]() ):
):
![]()
С учетом этого, общая фенотипическая варианса может быть представлена в следующем виде:
![]()