

1.3. Биологические мембраны и ионные каналы
Биологические мембраны – это функционально активные структуры клеток, ограничивающие цитоплазму и большинство внутриклеточных структур; образуют единую внутриклеточную систему канальцев, складок и замкнутых полостей.
Структурная основа мембраны – двойной слой фосфолипидов, в который встроены мембранные белки. Толщина клеточных мембран 6-12 нм. Молекулы липидов амфотерны. Своими гидрофильными частями они обращены в сторону водной среды (межклеточная жидкость и цитоплазма), гидрофобные части молекул направлены внутрь фосфолипидного бислоя. Такая структура идеально подходит для раздела внеклеточной и внутриклеточной фаз.
Белки, интегрированные в двойной слой фосфолипидов своими полярными участками, образуют гидрофильную поверхность в водной фазе. Они выполняют различные функции: рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул.
Большинство наших знаний об устройстве ионного канала, является результатом функциональной реконструкции. Каждый канал имеет устье, селективный фильтр, ворота и механизм управления воротами.
Часть каналов являются электроуправляемыми, т.е. управляются за счет разности потенциалов на мембране (потенциал-зависимые ионные каналы). Для этого рядом с каналом имеется электрический сенсор, который в зависимости от величины мембранного потенциала либо открывает ворота каналов, либо держит их закрытыми.
Второй вариант ионных каналов – рецептороуправляемые. Ворота управляются за счет рецептора, расположенного на поверхности мембраны (открываются при взаимодействии медиатора с рецептором). В некоторых рецептороуправляемых каналах между рецептором и воротным механизмом имеется промежуточная стадия (посредник типа цАМФ, протеинкиназы и т.д.)
Ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
Селективность – или избирательность канала обеспечивается его особой белковой структурой, геометрией канала.
Например, диаметр иона натрия – 0,19 нм, вместе с гидратной оболочкой он становится около 0,3 нм. Устье натриевого канала 0,3 – 0,5 нм. Чтобы пройти через канал (особенно через селективный фильтр), ион натрия или другой ион должен освободиться от гидратной оболочки и только в «голом» виде может пройти через канал. Слишком большой ион не может войти в устье, слишком маленький не способен отдать гидратную оболочку в селективном фильтре, поэтому не может выскочить из канала.
Натриевые каналы (рис. 6) имеют ворота 2-х типов – активационные (m-ворота) и инактивационные (h-ворота). В условиях покоя активационные ворота закрыты, но готовы в любую минуту открыться, а инактивационные – открыты. При снижении МП (деполяризация до 60 мВ) активационные ворота открываются и впускают ионы натрия в клетку, но вскоре начинают закрываться инактивационные ворота (происходит инактивация натриевых каналов). Некоторое время спустя закрываются активационные ворота, открываются инактивационные, и канал готов к новому циклу. Канал блокируется тетродотоксином, местными анестетиками (новокаин и др.).
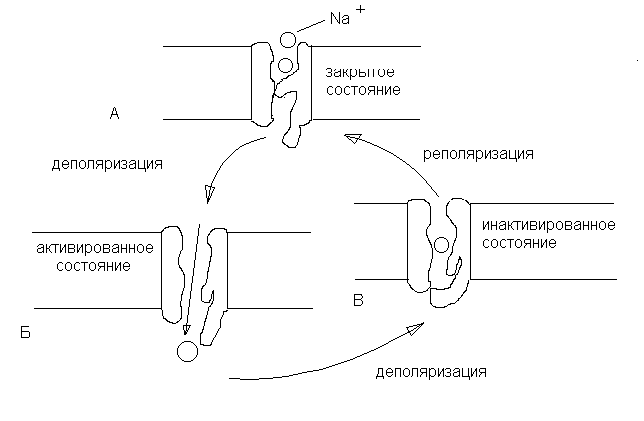
Рис. 6. Работа натриевых каналов и «воротных» механизмов.
А – в покое m-ворота закрыты; Б – при возбужденииm-ворота открыты; В – закрытиеh-ворот (инактивация) при деполяризации.
Калиевые каналы тоже достаточно селективны – в основном пропускают ионы калия. Блокируются тетраэтиламмонием. Процессы инактивации у них выражены слабо. Зато имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-калльцийзависимых каналов ускоряет реполяризацию (восстановление МП покоя).
Кальциевые каналы. Входящий кальциевый ток недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Кальций выступает в роли вторичного
посредника (мессенджера). Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, напр., входящим натриевым током. Инактивация кальциевых каналов происходит при повышении внутриклеточной концентрации свободного кальция. Однако белки цитоплазмы связывают кальций, что позволяет некоторое время поддерживать стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Блокируются кальциевые каналы ионами марганца, никеля, кадмия (2-хвалентные ионы), а также лекарственными веществами (верапамил).
Различают пассивный (без затрат энергии) и активный (энергозависимый) транспорт ионов через мембраны.
Пассивный идет за счет простой и облегченной диффузии.
Простая диффузия идет в соответствии с законом Фика – по химическому, электрохимическому или осмотическому градиенту. Напр., в клетке натрия 14 ммоль, а в среде 140 ммоль, в этом случае пассивный поток должен быть направлен в клетку.
Для пассивной простой диффузии вещество должно быть жирорастворимым. Гидрофильные вещества в фосфолипидном бислое мембраны пройти не могут.
Облегченная диффузия происходит или при наличии специализированных каналов или с участием переносчиков, которые специфически связываются с переносимой молекулой, а затем способствуют ее переносу по градиенту концентрации.
Активный транспорт ионов насосами клеточных мембран обеспечивает поддержание ионных градиентов по обе стороны мембраны. Энергия затрачивается на перенос данного вещества против градиента его концентрации.
Доказано участие в активном транспорте ионов специализированных ферментных систем – АТФ-аз, которые осуществляют гидролиз АТФ. Различают:
Натрий–калиевая–АТФ–аза («натриевый насос») обнаружена в мембранах клеток всех животных, растений и микроорганизмов. Это мембранный белок, имеющий два центра связывания ионов. Один из них (натриевый) расположен на внутренней поверхности клеточной мембраны, второй (калиевый) – на ее внешней поверхности. Специфическим ингибитором фермента является сердечный гликозид – строфантин (уабаин), блокирующий работу натриевого насоса. Гидролиз одной молекулы АТФ сопровождается выведением из клетки трех ионов натрия и закачиванием в клетку двух ионов калия. При увеличении количества ионов калия во внеклеточной среде или ионов натрия внутри клетки работа насоса усиливается.
Кальциевая–АТФ-аза («кальциевый насос») наиболее широко распространена в мембранах саркоплазматического ретикулума мышечных клеток.
Протонная–АТФ-аза («протонный насос») – в мембранах митохондрий.