

1
1. Предмет и задачи биологической химии. Биохимия как молекулярный уровень изучения структурной организации, анаболизма и катаболизма живой материи. Значение биохимии в подготовке врача.
Биологическая химия (биохимия) — это наука, изучающая химический состав живых организмов, превращения веществ и энергии, лежащей в основе их жизнедеятельности. Совокупность этих превращений составляет биологический обмен веществ, который является основой той формы движения материи, которую мы называем жизнью.
Живые организмы обладают необычными свойствами, отсутствующими в скоплении неживых молекул. К ним относятся следующие свойства: 1.1 Сложность и высокая степень организованности. Живые организмы представлены миллионами разных видов. 1.2 Любая составная часть организма имеет специальное назначение и выполняется строго определенную функцию. Это относится даже к индивидуальным химическим соединениям (липиды, белки и т.п.). 1.3Способность извлекать, преобразовывать и использовать энергию окружающей их среды – либо в форме органических питательных веществ, либо в виде энергии солнечного излучения
Обмен веществ слагается из множества отдельных химических реакций, протекающих в живом организме и теснейшим образом связанных друг с другом. Данные экспериментальной биохимии свидетельствуют о взаимосвязи и неразрывности процесса поглощения и усвоения питательных
веществ –ассимиляции и процесса |
их разложения |
и выделения |
– диссимиляции. |
Сопряженность и взаимосвязь |
отдельных реакций, |
происходящих при |
ассимиляции и |
диссимиляции питательных веществ в организме, проявляется также в сопряженности превращений
энергии, происходящих в течение всей жизни организма. |
|
|
|
|
||||
1.4 |
Способность |
к |
|
точному |
самовоспроизведению. |
|||
|
Цель биохимии состоит в том, чтобы понять, каким образом взаимодействия биомолекул друг с |
|||||||
другом порождают описанные выше особенности живого состояния. |
|
|
||||||
Биохимию |
|
|
разделяют |
|
|
на: |
||
3.1 |
Статическую, |
|
изучающую |
химический |
состав |
живой |
материи; |
|
3.2 |
Динамическую, |
изучающую |
процессы |
обмена |
веществ |
в |
организме; |
|
3.3 Функциональную, изучающую процессы, лежащие в основе определенных проявлений жизнедеятельности.
Первая часть обычно именуется органической химией и излагается в специальном курсе, вторая и третья части являются собственно биохимией.
Биологическая химия изучает молекулярные процессы, лежащие в основе развития и функционирования организмов. Биохимия использует методы «молекулярных» наук — химии, физической химии, молекулярной физики, и в этом отношении биохимия сама является молекулярной наукой.
Однако главные конечные задачи биохимии лежат в области биологии: она изучает закономерности биологической, а не химической формы движения материи. С другой стороны, «молекулярные изобретения» природы, открываемые биохимиками, находят применение в небиологических отраслях знания и в промышленности (молекулярная бионика, биотехнология). В таких случаях биохимия выступает в роли метода, а предметом исследований и разработок являются проблемы, выходящие за пределы биологии.
Место биохимии как молекулярного уровня биологических исследований. Уровни исследования являются отражением уровней структурной организации биологических систем, образующих иерархический ряд от наиболее простых систем (молекулы организмов, молекулярный уровень) до предельно сложной земной биологической системы (биосферный уровень). Действительные связи между отраслями биологии гораздо сложнее, чем можно представить с помощью таких простых схем. В частности, каждый более простой уровень организации живых систем (и, соответственно, уровень их исследования) является частью более сложных уровней. Самый первый уровень — молекулярный — уникален в том отношении, что он является составной частью систем всех других уровней биологии. Соответственно этому выделяют такие разделы биохимии, как, например, молекулярная генетика, биохимическая экология. Высший уровень — биосферный — включает в себя все другие уровни.
2
Значение биохимических исследований.
Из определения биологической химии вытекает, что это химия живых существ. Живая система от неживой отличается обменом веществ и энергии (метаболизмом).
В результате обмена веществ (метаболизма) в биологические внутренние среды нашего организма поступает большое количество продуктов обмена веществ (метаболитов), содержание которых у здорового человека варьирует незначительно и составляет гомеостаз внутренних сред организма (кровь, сыворотка, спинномозговая жидкость, моча, пищеварительные соки и др.).
Практически любое заболевание начинается с повреждения (нарушения) одной реакции в метаболизме клетки, а затем оно распространяется на ткань, орган и целый организм. Нарушение метаболизма ведет к нарушению гомеостаза в биологических жидкостях организма человека, что сопровождается изменением биохимических показателей.
Большое значение клинико-биохимических методов исследования биологических жидкостей велико в медицине и важно для подготовки медицинских лабораторных техников. Достаточно напомнить, что только в крови человека можно определить современными методами биохимических исследований около 1000 показателей метаболизма.
Биохимические показатели биологических сред организма человека широко используются при:
1.постановке диагноза заболевания, особенно дифференциального диагноза;
2.выборе метода лечения;
3.контроле за правильностью назначенного лечения; 4.результаты биохимических анализов служат одним из критериев излеченности патологического процесса; 5.скрининге (выявлении болезни на доклинической стадии);
6.мониторинге (контроле за течением заболевания и результатом лечения); 7. прогнозе (информации о возможном исходе заболевания).
Бурный рост биохимии привел к тому, что она подразделилась на разные отрасли: клиническую биохимию, молекулярную биохимию, биохимию спорта и биохимию человека.
В процессе освоения дисциплины «Основы биохимии с методами клинико-биохимических исследований» перед нами встают задачи медицинской биохимии, заключающиеся в изучении:
1.строения и функций биомолекул, входящих в состав тканей организма.
2.механизмов:
·поступления пластических и биологически активных веществ во внутреннюю среду организма;
·превращения поступивших мономеров в биополимеры, специфичные для данного организма;
·высвобождения, накопления и использования энергии в клетке;
·образования и выведения конечных продуктов распада веществ в организме;
·воспроизведения и передачи наследственных признаков организма;
·регуляции всех перечисленных процессов.
Основное внимание нашего курса будет уделяться изучению методов клинико-биохимических исследований, которые состоят из этапов.
2 Аминокислоты, входящие в состав белков, их строение и свойства. Биологическая роль аминокислот. Пептиды.
Белки - полимерные молекулы, в которых мономерами служат аминокислоты. В белках человека встречают только 20-АК.
А. Строение и свойства аминокислот
1. Общие структурные особенности аминокислот, входящих в состав белков
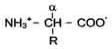
Общая структурная особенность АК - наличие амино- и карбоксильной групп, соединённых с одним и тем же углеродным атомом. R - радикал аминокислот - в простейшем случае представлен атомом водорода (глицин), но может иметь и более сложное строение.
3
Вводных растворах при нейтральном значении рН - АК существуют в виде биполярных ионов.
Вотличие от 19 остальных - АК, пролин - Иминокислота, радикал которой связан как с углеродным атомом, так и с аминогруппой, в результате чего молекула приобретает циклическую структуру.
19 из 20 АК содержат в α-положении асимметричный атом углерода, с которым связаны 4 разные замещающие группы. В результате эти АК в природе могут находиться в двух разных изомерных формах - L и D. Исключение составляет глицин, который не имеет асимметричного α-углеродного атома, так как его радикал представлен только атомом водорода. В составе белков присутствуют только L-изомеры аминокислот.
Чистые L- или D-стереоизомеры могут за длительный срок самопроизвольно и неферментатив-но превращаться в эквимолярную смесь L- и D-изомеров. Этот процесс называют рацемизацией. Рацемизация каждой L-аминокислоты при данной температуре идёт с определённой скоростью. Это обстоятельство можно использовать для установления возраста людей и животных. Так, в твёрдой эмали зубов имеется белок дентин, в котором L-аспартат переходит в D-изомер при температуре тела человека со скоростью 0,01% в год. В период формирования зубов в дентине содержится только L- изомер, поэтому по содержанию D-аспартата можно рассчитать возраст обследуемого.
Все 20 АК в организме человека различаются по строению, размерам и физико-химическим свойствам радикалов, присоединённых к α-углеродному атому.
2. Классификация аминокислот по химическому строению радикалов
По химическому строению АК можно разделить на алифатические, ароматические и гетероциклические.
Всоставе алифатических радикалов могут находиться функциональные группы, придающие им специфические свойства: карбоксильная (-СООН), амино (-NH2), тиольная (-SH), амидная (-CO-NH2),
гидроксильная (-ОН) и гуанидиновая 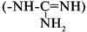 группы.
группы.
Названия аминокислот можно построить по заместительной номенклатуре, но обычно используют тривиальные названия.
3. Классификация аминокислот по растворимости их радикалов в воде
АК с неполярными R: радикалы, имеющие алифатические углеводородные цепи (радикалы ала, вал, лей, изо, про и мет) и ароматические кольца (радикалы фен и три).
АК с полярными незаряженными R: эти радикалы лучше, чем гидрофобные радикалы, растворяются в воде, т.к. в их состав входят полярные функциональные группы, образующие водородные связи с водой. К ним относят сер, тре и тир, имеющие гидроксильные группы, асн и глн, содержащие амидные группы, и цис с его тиольной группой.
Цистеин и тирозин содержат соответственно тиольную и гидроксильную группы, способные к диссоциации с образованием Н+, но при рН около 7,0, поддерживаемого в клетках, эти группы практически не диссоциируют.
АК с полярными отрицательно заряженными R: относят асн и глн аминокислоты, имеющие в радикале дополнительную карбоксильную группу, при рН около 7,0 диссоциирующую с образованием СОО- и Н+. Следовательно, радикалы данных аминокислот - анионы. Ионизированные формы глутаминовой и аспарагиновой кислот называют соответственно глутаматом и аспартатом.
АК с полярными положительно заряженными R:
α-Аминокислоты могут ковалентно связываться друг с другом с помощью пептидных связей. Пептидная связь образуется между α-карбоксильной группой одной аминокислоты и α-аминогруппой другой, т.е. является амидной связью. При этом происходит отщепление молекулы воды.
1. Строение пептида. Количество аминокислот в составе пептидов может сильно варьировать. Пептиды, содержащие до 10 аминокислот, называют олигопептиды. Часто в названии таких молекул указывают количество входящих в состав олигопептида аминокислот: трипептид, пентапептид, окгапептид и т.д.
Пептиды, содержащие более 10 аминокислот, называют "полипептиды", а полипептиды, состоящие из более чем 50 аминокислотных остатков, обычно называют белками. Однако эти названия условны, так как термин "белок" часто употребляют для обозначения полипептида, содержащего менее 50 аминокислотных остатков. Например, гормон глюкагон, состоящий из 29 аминокислот, называют белковым гормоном.
4
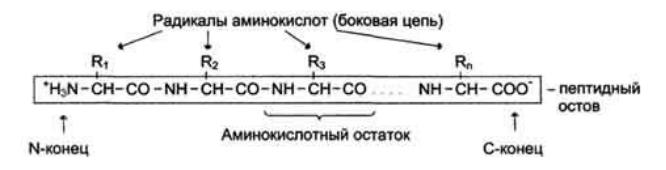
Мономеры аминокислот, входящих в состав белков, называют "аминокислотные остатки". Аминокислотный остаток, имеющий свободную аминогруппу, называется N-концевым и пишется слева, а имеющий свободную α-карбоксильную группу - С-концевым и пишется справа. Пептиды пишутся и читаются с N-конца. Цепь повторяющихся атомов в полипептидной цепи -NH- CH-CO-носит название "пептидный остов".
При названии полипептида к сокращённому названию аминокислотных остатков добавляют суффикс - ил, за исключением С-концевой аминокислоты. Например, тетрапептид Сер-Гли-Про-Ала читается как серилглицилпролилаланин.
Пептидная связь, образуемая иминогруппой пролина, отличается от других пептидных связей, так как атом азота пептидной группы связан не с водородом, а с радикалом.
Пептиды различаются по аминокислотному составу, количеству и порядку соединения аминокислот
3 Первичная структура белков. Пептидная связь, ее характеристика (прочность, кратность, компланарность, цис- ,транс- изомерия). Значение первичной структуры для нормального функционирования белков (на примере гемоглобина S).
Первичная структура — понятие, обозначающее последовательность аминокислотных остатков в белке Пептидная связь — основной вид связи, определяющий первичную структуру Возможно и присутствие дисульфидных связей между двумя остатками цистеина в одной полипептидной цепи с образованием цистина Такая же связь (дисульфидный мостик) может возникать и между остатками цистеина, принадлежащими разным полипептидным цепям в белковой молекуле, сополимерном образовании.
Аминокислотные остатки в пептидной цепи белков чередуются не случайным образом, а расположены в определённом порядке. Линейную последовательность аминокислотных остатков в полипептидной цепи называют "первичная структура белка".
Первичная структура каждого индивидуального белка закодирована в участке ДНК, называемом геном. В процессе синтеза белка информация, находящаяся в гене, сначала переписывается на мРНК, а затем, используя мРНК в качестве матрицы, на рибосоме происходит сборка первичной структуры белка.
Каждый из 50 000 индивидуальных белков организма человека имеет уникальную для данного белка первичную структуру. Все молекулы данного индивидуального белка имеют одинаковое чередование аминокислотных остатков в белке, что в первую очередь отличает данный индивидуальный белок от любого другого
2.Характеристика пептидной связи
Пептидная связь имеет характеристику частично двойной связи, поэтому она короче, чем остальные связи пептидного остова, и вследствие этого мало подвижна. Электронное строение пептидной связи определяет плоскую жёсткую структуру пептидной группы. Плоскости пептидных групп расположены под углом друг к другу.
5
Связь между α-углеродным атомом и α-аминогруппой или α-карбоксильной группой способна к свободным вращениям (хотя ограничена размером и характером радикалов), что позволяет полипептидной цепи принимать различные конфигурации.
Пептидные связи обычно расположены в транс-конфигурации, т.е. α-углеродные атомы располагаются по разные стороны от пептидной связи. В результате боковые радикалы аминокислот находятся на наиболее удалённом расстоянии друг от друга в пространстве.
Пептидные связи очень прочны и самопроизвольно не разрываются при нормальных условиях, существующих в клетках (нейтральная среда, температура тела). В лабораторных условиях гидролиз пептидных связей белков проводят в запаянной ампуле с концентрированной (6 моль/л) соляной кислотой, при температуре более 105 °С, причём полный гидролиз белка до свободных аминокислот проходит примерно за сутки.
Вживых организмах пептидные связи в белках разрываются с помощью специальных протеолитических ферментов (от англ, protein - белок, lysis - разрушение), называемых также протеазами, или пептидгидролазами.
Для обнаружения в растворе белков и пептидов, а также для их количественного определения используют биуретовую реакцию (положительный результат для веществ, содержащих в своём составе не менее двух пептидных связей).
Замена аминокислоты на поверхности гемоглобина А
Вмолекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 β-цепи, в которых глутамат, высокополярная отрицательно заряженная аминокислота в положении 6 была заменена валином, содержащим гидрофобный радикал.
Вдезоксигемоглобине S имеется участок, комплементарный другому участку таких же молекул, содержащему изменённую аминокислоту. В результате молекулы дезоксигемоглобина начинают "слипаться", образуя удлинённые фибриллярные агрегаты, деформирующие эритроцит и приводящие к образованию аномальных эритроцитов в виде серпа.
Воксигемоглобине S комплементарный участок "замаскирован" в результате изменения конформации белка. Недоступность участка препятствует соединению молекул оксигемоглобина S друг с другом. Следовательно, образованию агрегатов HbS способствуют условия, повышающие концентрацию дезоксигемоглобина в клетках (физическая работа, гипоксия, уменьшение рН, условия высокогорья, полёт на самолёте).
Так как "серповидные" эритроциты плохо проходят через капилляры тканей, они часто закупоривают сосуды и создают тем самым локальную гипоксию. Это повышает концентрацию дезоксигемоглобина S в эритроцитах, скорость образования агрегатов гемоглобина S и ещё большую деформацию
эритроцитов. Нарушение доставки О2 в ткани вызывает боли "даже некроз клеток в данной области. Серповидно-клеточная анемия - гомозиготное рецессивное заболевание; проявляется только в том случае, когда от обоих родителей наследуются 2 мутантных гена β-цепей глобина. После рождения ребёнка болезнь не проявляется до тех пор, пока значительные количества HbF не заместятся на HbS. У больных выявляют клинические симптомы, характерные для анемии: головокружение и головные боли, одышка, учащённое сердцебиение, боли в конечностях, повышенную восприимчивость к инфекционным заболеваниям.
Гетерозиготные индивидуумы, имеющие один нормальный ген НЬА, а другой ген HbS, в крови имеют лишь следовые количества серповидных клеток и нормальную продолжительность жизни; клинические симптомы болезни у них обычно не проявляются.
Для диагностики наличия HbS в эритроцитах человека используют метод электрофореза, основанного на движении заряженных белков в электрическом поле. Так как в HbS отрицательно заряженные группы глутамата в β-цепях заменены незаряженным валином, HbS в щелочной среде будет двигаться медленнее, чем НЬА.
Серповидно-клеточная: анемия - первый описанный пример молекулярной болезни.
Почти все встречающиеся замены аминокислот на поверхности молекулы гемоглобина безвредны. Гемоглобин S - редкое исключение.
4. Вторичная структура белков. Связи стабилизирующие вторичную структуру.
6
Вторичная структура белков - пространственная структура, образующаяся в результате взаимодействий между функциональными группами, входящими в состав пептидного остова. При этом пептидные цепи могут приобретать регулярные структуры двух типов: α-спираль и β-структура.
α-спираль
Пептидный остов закручивается в виде спирали за счёт образования водородных связей между атомами кислорода карбонильных групп и атомами азота аминогрупп, входящих в состав пептидных групп через 4 аминокислотных остатка. Водородные связи ориентированы вдоль оси спирали. На один виток α-спирали приходится 3,6 аминокислотных остатка.
Вобразовании водородных связей участвуют практически все атомы кислорода и водорода пептидных групп. В результате α-спираль "стягивается" множеством водородных связей. Несмотря на то, что данные связи относят к разряду слабых, их количество обеспечивает максимально возможную стабильность α -спирали. Так как все гидрофильные группы пептидного остова обычно участвуют в образовании водородных связей, гидрофильность (т.е. способность образовывать водородные связи с водой) α-спиралей уменьшается, а их гидрофобность увеличивается.
α-Спиральная структура - наиболее устойчивая конформация пептидного остова, отвечающая минимуму свободной энергии. В результате образования α-спиралей полипептидная цепь укорачивается, но если создать условия для разрыва водородных связей, полипептидная цепь вновь удлинится.
Радикалы аминокислот находятся на наружной стороне α-спирали и направлены от пептидного остова в стороны. Они не участвуют в образовании водородных связей, характерных для вторичной структуры, но некоторые из них могут нарушать формирование α-спирали. К ним относят:
-пролин. Его атом азота входит в состав жёсткого кольца, что исключает возможность вращения вокруг -N-CH- связи. Кроме того, у атома азота пролита, образующего пептидную связь с другой аминокислотой, нет атома водорода. В результате пролин не способен образовать водородную связь в данном месте пептидного остова, и α-спиральная структура нарушается. Обычно в этом месте пептидной цепи возникает петля или изгиб; -участки, где последовательно расположены несколько одинаково заряженных радикалов, между
которыми возникают электростатические силы отталкивания; -участки с близко расположенными объёмными радикалами, механически нарушающими формирование α-спирали, например метионин, триптофан.
β-Структура
β -Структура формируется за счёт образования множества водородных связей между атомами пептидных групп линейных областей одной полипептидной цепи, делающей изгибы, или между разными полипептидными цепями, β -Структура образует фигуру, подобную листу, сложенному "гармошкой", - β -складчатый слой.
Когда водородные связи образуются между атомами пептидного остова различных полипептидных цепей, их называют межцепочечными связями. Водородные связи, возникающие между линейными участками внутри одной полипептидной цепи, называют внутрицепочечными. В β -структурах водородные связи расположены перпендикулярно полипептидной цепи.
Если связанные полипептидные цепи направлены противоположно, возникает антипараллельная β - структура, если же N- и С-концы полипептидных цепей совпадают, образуется структура параллельного ?-складчатого слоя.
Вотличие от β-спиралей, разрыв водородных связей, формирующих β-структуры, не вызывает удлинения данных участков полипептидных цепей.
Как β-спираль, так и β-структуры обнаружены в глобулярных и фибриллярных белках.
Нерегулярные вторичные структуры
В белках отмечают области с нерегулярной вторичной структурой, которые часто называют беспорядочными клубками. Они представлены петлеобразными и кольцеобразными структурами, имеющими меньшую регулярность укладки, чем описанные выше α-спиралъ и β-структура. Однако и они не так сильно варьируют от одной молекулы белка к другой. В каждом индивидуальном белке они имеют свою фиксированную конформацию, определяемую аминокислотным составом данного участка цепи и окружающих его участков.
7
Термином "беспорядочный клубок" также часто называют денатурированный белок, образовавшийся после разрыва слабых внутримолекулярных связей и потерявший свою упорядоченную структуру.
Содержание разных типов вторичных структур в белках
Содержание рассмотренных выше типов вторичных структур в разных белках неодинаково. По наличию α-спиралей и β-структур глобулярные белки можно разделить на 4 категории.
К 1ой категории относят белки, в структуре которых обнаружены только α-спирали. К ним принадлежат такие белки, как миоглобин и гемоглобин .
Ко 2ой категории относят белки с α-спиралями и β-структурами, иногда образующими однотипные сочетания, встречающиеся в разных индивидуальных белках.
Характерные сочетания α-спиралей и β-структур, обнаруженные во многих ферментах, можно рассмотреть на примере строения доменов лактатдегидрогеназы (ДЦГ) и фосфоглицераткиназы (ФГК). Домен - участок полипептидной цепи, который самостоятельно от других участков той же цепи образует структуру, во многом напоминающую глобулярный белок.
Водном из доменов лактатдегидрогеназы в центре расположены β-структуры полипептидной цепи в виде скрученного листа, и каждая β-структура связана с α-спиральным участком, находящимся на поверхности молекулы.
Втретью категорию включены белки, имеющие только β-структуры. Такие структуры обнаружены в иммуноглобулинах, в ферменте супероксиддисмутазе.
Вчетвёртую категорию включены белки, имеющие в своём составе лишь незначительное количество регулярных вторичных структур.
5 Третичная структура белков. Типы химических связей, участвующих в формировании третичной структуры. Супервторичная структура. Доменная структура и ее роль в функционировании белков. Роль шаперонов (белки теплового шока) в формировании третичной структуры белков in vivo.
Третичная структура белков - трёхмерная пространственная структура, образующаяся за счёт взаимодействий между радикалами аминокислот, которые могут располагаться на значительном расстоянии друг от друга в полипептидной цепи.
Связи, участвующие в формировании третичной структуры белков
Гидрофобные взаимодействия
При укладке полипептидная цепь белка стремится принять энергетически выгодную форму, характеризующуюся минимумом свободной энергии. Поэтому гидрофобные радикалы аминокислот стремятся к объединению внутри глобулярной структуры растворимых в воде белков. Между ними возникают так называемые гидрофобные взаимодействия, а также силы ван дер Ваальса между близко
прилегающими друг |
к другу атомами. |
В результате внутри |
белковой глобулы |
формируется гидрофобное |
ядро. Гидрофильные |
группы пептидного остова |
при формировании |
вторичной структуры образуют множество водородных связей, благодаря чему исключается связывание с ними воды и разрушение внутренней, плотной структуры белка.
Ионные и водородные связи. Гидрофильные радикалы аминокислот стремятся образовать водородные связи с водой и поэтому в основном располагаются на поверхности белковой молекулы.
Все гидрофильные группы радикалов аминокислот, оказавшиеся внутри гидрофобного ядра, взаимодействуют друг с другом с помощью ионных и водородных связей.
Ионные связи могут возникать между отрицательно заряженными (анионными) карбоксильными группами радикалов аспарагиновой и глутаминовой кислот и положительно заряженными (катионными) группами радикалов лизина, аргинина или гистидина.
Водородные связи возникают между гидрофильными незаряженными группами (такими как -ОН, - CONH2, SH-группы) и любыми другими гидрофильными группами. Белки, функционирующие в неполярном (липидном) окружении, например белки мембран, имеют обратное устройство: гидрофильные радикалы аминокислот расположены внутри белка, в то время как гидрофобные аминокислоты локализованы на поверхности молекулы и контактируют с неполярным окружением. В каждом случае радикалы аминокислот занимают наиболее выгодное биоэнергетическое положение.
Ковалентные связи. |
Третичную структуру |
некоторых белков стабилизируют дисульфидные |
связи, образующиеся |
за счёт взаимодействия |
SH-групп двух остатков цистеина. Эти два остатка |
8
цистеина могут находиться далеко друг от друга в линейной первичной структуре белка, но при формировании третичной структуры они сближаются и образуют прочное ковалентное связывание радикалов.
Большинство внутриклеточных белков лишено дисульфидных связей. Однако такие связи распространены в белках, секретируемых клеткой во внеклеточное пространство. Полагают, что эти ковалентные связи стабилизируют кон-формацию белков вне клетки и предотвращают их денатурацию. К таким белкам относят гормон инсулин и иммуноглобулины.
Все белки с одинаковой первичной структурой, находящиеся в одинаковых условиях, приобретают одинаковую, характерную для данного индивидуального белка конформацию, определяющую его специфическую функцию. Функционально активную конформацию белка называют "нативная структура".
Супервторичная структура белков
Супервторичная структура формируется за счёт межрадикальных взаимодействий.
Определённые характерные сочетания α-спиралей и β-структур часто обозначают как "структурные мотивы". Они имеют специфические названия: "α-спираль-поворот- α-спираль", "структура β- бочонка", "лейциновая застёжка-молния", "цинковый палец" и др. Специфическое пространственное расположение α-спиралей и β-структур формируется за счёт межрадикальных взаимодействий.
1.Супервторичная структура типа β-бочонка. Такая структура действительно напоминает бочонок, где каждая β-структура расположена внутри и связана с α-спиральным участком полипептидной цепи, находящимся на поверхности молекулы.
Супервторичную структуру в виде β-бочонка имеют некоторые ферменты, например триозофосфатизомераза и один домен пируваткиназы.
2.Структурный мотив "α-спираль-поворот-α-спиралъ". Этот "структурный мотив" обнаружен во многих ДНК-связывающих белках. Двухспиральная структура ДНК имеет две бороздки - большую и малую. Большая бороздка хорошо приспособлена для связывания белков, имеющих небольшие спиральные участки. В данный структурный мотив входят две α-спирали: одна более короткая, другая более длинная, которые соединены поворотом полипептидной цепи. Более короткая α-спираль располагается поперёк бороздки, а более длинная α-спираль - в большой бороздке, образуя нековалентные специфические связи радикалов аминокислот с нуклеотидами ДНК.
3.Супервторичная структура в виде "цинкового пальца". Этот вид супервторичной структуры также часто отмечают в ДНК-связывающих белках. "Цинковый палец" - фрагмент белка, содержащий около 20 аминокислотных остатков, в котором атом цинка связан с радикалами четырёх аминокислот: обычно с двумя остатками цистеина и двумя - гистидина. В некоторых случаях вместо остатков гистидина также находятся остатки цистеина.
Два близко лежащих остатка цистеина отделены от двух других остатков гистидина (или цистеина) аминокислотной последовательностью, состоящей примерно из 12 аминокислотных остатков. Этот участок белка образует α-спираль, которая может специфично связываться с регуляторными участками большой бороздки ДНК. Специфичность взаимодействия ДНК-связывающего белка с определённой областью ДНК зависит от последовательности аминокислотных остатков, расположенных в области "цинкового пальца".
4.Супервторичная структура в виде "лейциновой застёжки-молнии". Некоторые ДНК-
связывающие белки олигомерны, т.е. содержат в своём составе несколько полипептидных цепей. Кроме того, существуют белки, которые функционируют в комплексе с другими белками. Объединение протомеров или отдельных белков в комплексы иногда осуществляется с помощью структурных мотивов, называемых "лейциновая застёжка-молния". На поверхности каждой из 2х взаимодействующих полипептидных цепей или белков имеется α-спиральный участок, содержащий по крайней мере 4 остатка лейцина. Лейциновые остатки располагаются через каждые 6 аминокислот один от другого. Так как каждый виток α-спирали содержит 3,6 аминокислотных остатка, радикалы лейцина находятся на поверхности каждого второго витка.
Лейциновые остатки α-спирали одного белка могут взаимодействовать с лейциновыми остатками другого белка с помощью гидрофобных взаимодействий, соединяя их вместе.
Примером соединения белков с помощью "лейциновой застёжки-молнии" могут служить гистоны. Гистоны - ядерные белки, в состав которых входит большое количество положительно заряженных аминокислот - аргинина и лизина. Молекулы гистонов объединяются в комплексы, состоящие из 8
9
мономерных белков с помощью "лейциновых застёжек", несмотря на то, что все мономеры имеют сильный положительный заряд.
Доменная структура белков. Если полипептидная цепь белка содержит более 200 аминокислот, как правило, её пространственная структура сформирована в виде двух или более доменов. Домен - участок полипептидной цепи, который в процессе формирования пространственной структуры приобрёл независимо от других участков той же цепи конформацию глобулярного белка. Так, лёгкая цепь иммуноглобулина G состоит из двух доменов. В некоторых случаях доменами называют отдельные структурные участки полипептидной цепи.
Домены обычно можно выделить, действуя на белок протеолитическими ферментами, легко разрывающими пептидные связи на участке полипептидной цепи, расположенной между доменами. После этого некоторые домены могут сохранять свои биологические свойства.
Структура и функциональная роль шаперонов в фолдинге белков. В процессе синтеза полипептидных цепей, транспорта их через мембраны, при сборке олигомерных белков возникают промежуточные нестабильные конформации, склонные к агрегации. На вновь синтезированном полипептиде имеется множество гидрофобных радикалов, которые в трёхмерной структуре спрятаны внутри молекулы. Поэтому на время формирования нативной конформации реакционно-способные аминокислотные остатки одних белков должны быть отделены от таких же групп других белков.
Во всех известных организмах от прокариотов до высших эукариотов обнаружены белки, способные связываться с белками, находящимися в неустойчивом, склонном к агрегации состоянии. Они способны стабилизировать их конформацию, обеспечивая фолдинг белков. Эти белки получили название "шапероны".
1. Классификации шаперонов (Ш)
В соответствии с молекулярной массой все шапероны можно разделить на 6 основных групп: высокомолекулярные, с молекулярной массой от 100 до 110 кД; Ш-90 - с молекулярной массой от 83 до 90 кД; Ш-70 - с молекулярной массой от 66 до 78 кД;
Ш-60; Ш-40;
низкомолекулярные шапероны с молекулярной массой от 15 до 30 кД.
Среди шаперонов различают: конститутивные белки (высокий базальный синтез которых не зависит от стрессовых воздействий на клетки организма), и индуцибельные, синтез которых в нормальных условиях идёт слабо, но при стрессовых воздействиях на клетку резко увеличивается. Индуцибельные шапероны относят к "белкам теплового шока", быстрый синтез которых отмечают практически во всех клетках, которые подвергаются любым стрессовым воздействиям. Название "белки теплового шока" возникло в результате того, что впервые эти белки были обнаружены в клетках, которые подвергались воздействию высокой температуры.
2. Роль шаперонов в фолдинге белков
При синтезе белков N-концевая область полипептида синтезируется раньше, чем С-концевая область. Для формирования конформации белка нужна его полная аминокислотная последовательность. Поэтому в период синтеза белка на рибосоме защиту реакционно-способных радикалов (особенно гидрофобных) осуществляют Ш-70.
Ш-70 - высококонсервативный класс белков, который присутствует во всех отделах клетки: цитоплазме, ядре, ЭР, митохондриях. В области карбоксильного конца единственной полипептидной цепи шаперонов есть участок, образованный радикалами аминокислот в форме бороздки. Он способен взаимодействовать с участками белковых молекул и развёрнутых полипептидных цепей длиной в 7-9 аминокислот, обогащённых гидрофобными радикалами. В синтезирующейся полипептидной цепи такие участки встречают примерно через каждые 16 аминокислот.
Фолдинг многих высокомолекулярных белков, имеющих сложную конформацию (например, доменное строение), осуществляется в специальном пространстве, сформированном Ш-60. Ш-60 функционируют в виде олигомернoго комплекса, состоящего из 14 субъединиц.
Ш-60 образуют 2 кольца, каждое из которых состоит из 7 субъединиц, соединённых друг с другом. Субъединица Ш-60 состоит из 3 доменов: апикального (верхушечного), промежуточного и экваториального. Верхушечный домен имеет ряд гидрофобных остатков, обращённых в полость
10
кольца, сформированного субъединицами. Экваториальный домен имеет участок связывания с АТФ и обладает АТФ-азной активностью, т.е. способен гидролизовать АТФ до АДФ и Н3РО4.
Шапероновый комплекс имеет высокое сродство к белкам, на поверхности которых есть элементы, характерные для несвёрнутых молекул (прежде всего участки, обогащённые гидрофобными радикалами). Попадая в полость шаперонового комплекса, белок связывается с гидрофобными радикалами апикальных участков Ш-60. В специфической среде этой полости, в изоляции от других молекул клетки происходит перебор возможных конформации белка, пока не будет найдена единственная, энергетически наиболее выгодная конформация.
Высвобождение белка со сформированной нативной конформацией сопровождается гидролизом АТФ в экваториальном домене. Если белок не приобрёл нативной конформации, то он вступает в повторную связь с шапероновым комплексом. Такой шаперонзависимый фолдинг белков требует затрат большого количества энергии.
Таким образом, синтез и фолдинг белков протекают при участии разных групп шаперонов, препятствующих нежелательным взаимодействиям белков с другими молекулами клетки и сопровождающих их до окончательного формирования нативной структуры.
6 Активный центр белков и его специфическое взаимодействие с лигандом как основа биологической функции белков. Конформационная лабильность белков. Комплементарность взаимодействия белков с лигандами. Обратимость связывания.
Активный центр белков - определённый участок белковой молекулы, как правило, находящийся в её углублении ("кармане"), сформированный радикалами аминокислот, собранных на определённом пространственном участке при формировании третичной структуры и способный комплементарно связываться с лигандом. В линейной последовательности полипептидной цепи радикалы, формирующие активный центр, могут находиться на значительном расстоянии друг от друга.
Высокая специфичность связывания белка с лигандом обеспечивается комплементарностью структуры активного центра белка структуре лиганда.
Под комплементарностью понимают пространственное и химическое соответствие взаимодействующих молекул. Лиганд должен обладать способностью входить и пространственно совпадать с конформацией активного центра. Это совпадение может быть неполным, но благодаря конформационной лабильности белка активный центр способен к небольшим изменениям и "подгоняется" под лиганд. Кроме того, между функциональными группами лиганда и радикалами аминокислот, образующих активный центр, должны возникать связи, удерживающие лиганд в активном центре. Связи между лигандом и активным центром белка могут быть как нековалентными (ионными, водородными, гидрофобными), так и ковалентными.
Характеристика активного центра. Активный центр белка - относительно изолированный от окружающей белок среды участок, сформированный аминокислотными остатками. В этом участке каждый остаток благодаря своему индивидуальному размеру и функциональным группам формирует "рельеф" активного центра.
Объединение таких аминокислот в единый функциональный комплекс изменяет реакционную способность их радикалов, подобно тому, как меняется звучание музыкального инструмента в ансамбле. Поэтому аминокислотные остатки, входящие в состав активного центра, часто называют "ансамблем" аминокислот.
Уникальные свойства активного центра зависят не только от химических свойств формирующих его аминокислот, но и от их точной взаимной ориентации в пространстве. Поэтому даже незначительные нарушения общей конформации белка в результате точечных изменений его первичной структуры или условий окружающей среды могут привести к изменению химических и функциональных свойств радикалов, формирующих активный центр, нарушать связывание белка с лигандом и его функцию. При денатурации активный центр белков разрушается, и происходит утрата их биологической активности.
Часто активный центр формируется таким образом, что доступ воды к функциональным группам его радикалов ограничен, т.е. создаются условия для связывания лиганда с радикалами аминокислот.
В некоторых случаях лиганд присоединяется только к одному из атомов, обладающему определённой реакционной способностью, например присоединение О2 к железу миоглобина или гемоглобина.