

231
гена выживали при эпидемиях малярии, однако четверть их потомства погибала от серповидноклеточной анемии.
Талассемии - наследственные заболевания, обусловленные отсутствием или снижением скорости синтеза α- или β-цепей гемоглобина. В результате несбалансированного образования глобиновых цепей образуются тетрамеры гемоглобина, состоящие из одинаковых протомеров. Это приводит к нарушению основной функции гемоглобина - транспорту кислорода к тканям. Нарушение эритропоэза и ускоренный гемолиз эритроцитов и клеток-предшественников при талассемиях приводит к анемии.
При β-талассемии не синтезируются β-цепи гемоглобина. Это вызывает образование нестабильных тетрамеров, содержащих только α-цепи. При этом заболевании в костном мозге из-за преципитации нестабильных α-цепей усиливается разрушение эритробластов, а ускорение разрушения эритроцитов в циркулирующей крови приводит к внутрисосудистому гемолизу. Как известно, для образования фетального гемоглобина р-цепи не требуются (см. раздел 4), поэтому клинически β- талассемия не проявляется до рождения, после чего происходит переключение синтеза HbF на НBА.
В случае α-талассемии недостаток образования α-глобиновых цепей приводит к нарушению образования HbF у плода. Избыточные γ-цепи образуют тетрамеры, называемые гемоглобином Барта. Этот гемоглобин при физиологических условиях имеет повышенное сродство к кислороду и не проявляет кооперативных взаимодействий между протомерами. В результате гемоглобин Барта не обеспечивает развивающийся плод необходимым количеством кислорода, что приводит к тяжёлой гипоксии. При α-талассемии отмечают высокий процент внутриутробной гибели плода. Выжившие новорождённые при переключении с γ- на β-ген синтезируют β-тетрамеры или НBН, который, подобно гемоглобину Барта, имеет слишком высокое сродство к кислороду, менее стабилен, чем НBА и быстро разрушается. Это ведёт к развитию у больных тканевой гипоксии и к смерти вскоре после рождения.
Наследственный сфероцитоз. Причиной этой патологии чаще всего является дефект белков цитоскелета эритроцитов - спектрина или ан-кирина, которые обеспечивают поддержание двояковогнутой формы клетки и эластичности мембраны. Эритроциты приобретают шарообразную форму, что приводит к уменьшению площади их поверхности и снижению скорости газообмена. Потеря эластичности клеточной мембраны приводит к повышению хрупкости и травматичности клеток и, как следствие, к ускорению их разрушения в сосудистом русле и селезёнке. Заболевание сопровождается анемией и желтухой. Удаление селезёнки (спленэктомия) при наследственном сфероцитозе улучшает состояние больных, так как предотвращает разрушение сфероцитов в селезёнке.
Мегалобластная (макроцитарная) анемия развивается при дефиците фолиевой кислоты или витамина В12.
Фолиевая кислота в виде кофермента (Н4-фолата) участвует в синтезе нуклеотидов. Недостаток фолиевой кислоты приводит к снижению скорости синтеза ДНК в быстроделящихся клетках, и в первую очередь в предшественниках эритроцитов. Клетки дольше пребывают в интерфазе, синтезируя гемоглобин, и становятся крупнее. Кроме того, из-за недостатка нуклеотидов они реже делятся, и количество эритроцитов снижается, а крупные мегалобласты быстрее разрушаются. Всё это в конечном итоге приводит к развитию анемии.
Аналогичная симптоматика развивается при недостатке в организме витамина В12. Этот витамин участвует в переносе метальной группы с N5-метил-Н4-фолата на гомоцистеин с образованием метионина и Н4-фолата (см. раздел 10). Недостаточность витамина В12 приводит к накоплению N5- метил-Н4-фолата в клетках. Дефицит Н4-фолата приводит к нарушению деления клеток и развитию анемии.
232
97. Биосинтез гема. Схема процесса, химизм первых двух реакций, место протекания. Регуляция
активности ферментов АЛК-синтазы и АЛК-дегидратазы. Источники железа для синтеза гема, всасывание железа, транспорт в крови, депонирование.
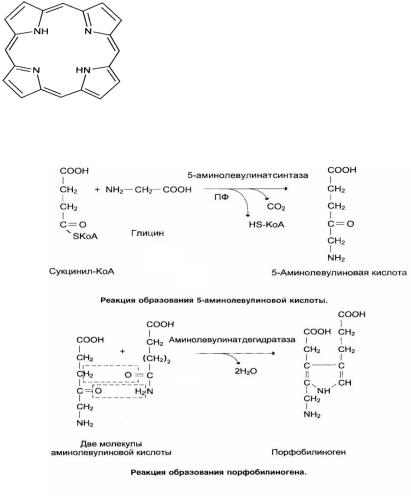
Гемоглобин – это сложный белок, простетической группой которого является гем. Основная роль гемоглобина – перенос кислорода и углекислого газа.
Гем – комплекс железа (Fe+2) c протопорфирином.
Биосинтез гема
Наиболее активно гем синтезируется в костном мозге и печени.
В костном мозге гем синтезируется для образования гемоглобина в ретикулоцитах, в печени – гем нужен для синтеза цитохрома P450.
Предшественниками синтеза гема являются глицин и сукцинил-КоА, поэтому путь синтеза гема называется глициносукцинатный цикл.
Первые две реакции приводят к образованию порфобилиногена. Далее идёт образование линейных тетрапирролов, которые завершается синтезом протопорфирина IX. Протопорфирин IX присоединяет Fe2+ с помощью фермента – феррохелатазы (гем-синтезы) и образуется гем.
Регуляция синтеза гема
Регуляторными ферментами являются АЛК (аминолевулинатсинтетаза) и аминолевулинатдегидрогеназа.
АЛК - синтаза является главным регуляторным ферментом, коферментом которого служит ПФ.
Аллостерическими ингибиторами АЛКсинтетазы служит гем и гемоглобин.
На уровне трансляции АЛК-синтазы регуляция осуществляется концентрацией железа.
В точке инициации мРНК АЛК-синтазы имеется шпилечная петля, которая называется железочувствительный элемент (IRE).
Регуляторный железосвязывающий белок при высокой концентрации железа связывается с железом и вызывает
снижение сродства этого белка к IRE – элементу мРНК АЛК-синтетазы и продолжение трансляции.
При низких концентрациях железа увеличивается сродство железосвязывающего белка к IREэлементу мРНК АЛК-синтетазы и трансляция аминолевупинатсинтетазы тормозится.
Недостаток ПФ и лекарства – аналоги ПФ снижают активность АЛК-синтетазы. Стероиды, барбитураты, сульфаниламиды, эстрогены – индукторы синтеза АЛК.

233
Нарушения синтеза гема – Порфирии
Порфирии – болезни, связанные с нарушением работы ферментов синтеза гема. Первичные порфирии обусловлены генетическими дефектами ферментов синтеза гема, вторичные связаны с нарушениями регуляции синтеза гема.
В зависимости от основной локализации патологического процесса различают печёночные и эритропоэтические наследственные порфирии. При этом эритропоэтические порфирии сопровождаются
накоплением порфиринов в нормобластах и эритроцитах, а печёночные – в гепатоцитах. Порфириногены не окрашены, но на свету они легко переходят в порфирины, которые проявляют красную флуоресценцию в ультрафиолетовых лучах.
Вкоже на солнце кислород реагирует с порфиринами и переходит в синглетное состояние. В этой форме он вызывает ПОЛ клеточных мембран и разрушение клеток, поэтому порфирии часто сопровождаются фотосенсибилизацией и изъязвлением открытых участков кожи.
Аминолевулинат и порфириногены являются нейротоксинами, что приводит к нейропсихическим расстройствам.
Приём лекарств – индукторов АЛК – синтетазы – сульфаниламидов, барбитуратов, диклофенака, вольтарена, стероидов, истогенов – может вызвать обострение порфирии.
Впериод полового созревания при повышении образования β-стероидов идёт индукция синтеза АЛК-синтетазы и проявляются симптомы порфирий.
Порфирии наблюдаются и при отравлении солями свинца, т.к. свинец ингибирует АЛКдегидратазу и феррохелатазу.
Обмен железа
Источником железа для синтеза гема служат пищевые продукты и железо, освобождающееся при распаде эритроцитов.
В гем-содержащих белках железо находится в составе гема.

234
Внегемовых железосодержащих белках железо непосредственно связывается с белком. К таким белкам относят трансферрин, ферритин, рибонуклеотидредуктазу, ксантиноксидазу и др.
Ворганизме взрослого человека содержится 3-4 г. железа.
Впище железо находится в окислённом состоянии (Fe+3). При кислом значении pH желудка железо выделяется из солей органических кислот.
Аскорбиновая кислота (Витамин С), содержащаяся в пище, восстанавливает железо и улучшает его всасывание и в клетки слизистой оболочки кишечника поступает железо в виде Fe+2.
Апоферритин в энтероцитах связывает железо и превращается в ферритин – запасную форму железа.
Железо по крови транспортируется трансферрином.
Трансферрин – гликопротеин, который синтезируется в печени и связывает только окисленное железо (Fe+3).
Поступающее в кровь железо окисляется ферментом феррооксидазой (церулоплазмином). Этом фермент содержит медь.
Трансферрин может соединиться с одним или двумя ионами Fe+3 одновременно с анионом , образуя комплекс трансферрин – 2 (Fe+3 - ).
Трансферрин взаимодействует со специфическими рецепторами клеток. В результате в цитозоле клетки образуется комплекс Ca+2 – кальмодулин – ПКС, который фосфорилирует рецептор трансферрина и вызывает образование эндосомы.
Вэндосоме создаётся кислая среда при действии АТФ – зависимого протонного насоса. При этом железо освобождается из трансферрина.
Ферритин – олигомерный белок, состоит из 24 протолиров и служит для запасания железа в клетке.
Вразных тканях существуют его изоформы.
Ферритин имеет полость, внутри которой может содержаться до 4500 ионов Fe+3.
Fe+2 окисляют в Fe+3 тяжёлые цепи ферритина. Комплекс железа в виде гидрофосфата хранится внутри сферы, образованной белковой частью молекулы.
Наибольшая часть ферритина содержится в печени, селезёнке и костном мозге.
98. Белки сыворотки крови, биологическая роль основных фракций белков, значение их определения для диагностики заболевания. Ферменты плазмы крови, энзимодиагностика. Количественное определение активности аминотрансфераз (АЛТ, АСТ).
В плазме крови содержится 7% всех белков организма при концентрации 60 - 80 г/л. Белки плазмы крови выполняют множество функций. Одна из них заключается в поддержании осмотического давления, так как белки связывают воду и удерживают её в кровеносном русле.
Белки плазмы образуют важнейшую буферную систему крови и поддерживают рН крови в пределах 7,37 - 7,43.
Альбумин, транстиретин, транскортин, трансферрин и некоторые другие белки (табл. 14-2) вьшолняют транспортную функцию.
235
Белки плазмы определяют вязкость крови и, следовательно, играют важную роль в гемодинамике кровеносной системы.
Белки плазмы крови являются резервом аминокислот для организма.
Иммуноглобулины, белки свёртывающей системы крови, α1-антитрипсин и белки системы комплемента осуществляют защитную функцию.
Методом электрофореза на ацетилцеллюлозе или геле агарозы белки плазмы крови можно разделить на альбумины (55-65%), α1-глобулины (2- 4%), α2 -глобулины (6-12%), β-глобулины (8-12%) и γ-глобулины (12-22%) (рис. 14-19).
Применение других сред для электрофоретического разделения белков позволяет обнаружить большее количество фракций. Например, при электрофорезе в полиакриламидном или крахмальном гелях в плазме крови выделяют 16-17 белковых фракций. Метод иммуноэлектрофореза, сочетающий электрофоретический и иммунологический способы анализа, позволяет разделить белки плазмы крови более чем на 30 фракций.
Большинство сывороточных белков синтезируется в печени, однако некоторые образуются и в других тканях. Например, γ-глобулины синтезируются В-лимфоцитами (см. раздел 4), пептидные гормоны в основном секретируют клетки эндокринных желёз, а пептидный гормон эритропоэтин - клетки почки.
Для многих белков плазмы, например альбумина, α1-антитрипсина, гаптоглобина, транс-феррина, церулоплазмина, α2-макроглобулина и иммуноглобулинов, характерен полиморфизм (см. раздел 4).
Почти все белки плазмы, за исключением альбумина, являются гликопротеинами. Олигосахариды присоединяются к белкам, образуя гликозидные связи с гидроксильной группой серина или треонина, или взаимодействуя с карбоксильной группой аспарагина. Концевой остаток олигосахаридов в большинстве случаев представляет собой N-ацетилнейраминовую кислоту, соединённую с галактозой. Фермент эндотелия сосудов нейраминидаза гидролизует связь между ними, и галактоза становится доступной для специфических рецепторов гепатоцитов. Путём эвддцитоза "состарившиеся" белки поступают в клетки печени, где разрушаются. Т1/2 белков плазмы крови составляет от нескольких часов до нескольких недель.
При ряде заболеваний происходит изменение соотношения распределения белковых фракций при электрофорезе по сравнению с нормой (рис. 14-20).
Такие изменения называют диспротеинемиями, однако их интерпретация часто имеет относительную диагностическую ценность. Например, характерное для нефротического синдрома снижение альбуминов, α1- и γ-глобулинов и увеличение α2- и β-глобулинов отмечают и при некоторых других заболеваниях, сопровождающихся потерей белков. При снижении гуморального иммунитета уменьшение фракции γ-глобулинов свидетельствует об уменьшении содержания основного компонента иммуноглобулинов - IgG, но не отражает динамику изменений IgA и IgM.
Содержание некоторых белков в плазме крови может резко увеличиваться при острых воспалительных процессах и некоторых других патологических состояниях (травмы, ожоги, инфаркт миокарда). Такие белки называют белками острой фазы, так как они принимают участие в развитии воспалительной реакции организма. Основной индуктор синтеза большинства белков острой фазы в гепатоцитах - полипептид интерлейкин-1, освобождающийся из мононуклеарных фагоцитов. К белкам острой фазы относят С-реактивный белок, называемый так, потому что он взаимодействует с С- полисахари-дом пневмококков, α1-антитрипсин, гаптоглобин, кислый гликопротеин, фибриноген. Известно, что С-реактивный белок может стимулировать

236
Рис. 14-19. Электрофореграмма (А) и денситограмма (Б) белков сыворотки крови.
Рис. 14-20. Протеинограммы белков сыворотки крови. а - в норме; б - при нефротическом синдроме; в - при гипогаммаглобулинемии; г - при циррозе печени; д - при недостатке α1- антитрипсина; е - при диффузной гипергаммаглобулинемии.
систему комплемента, и его концентрация в крови, например, при обострении ревматоидного артрита может возрастать в 30 раз по сравнению с нормой. Белок плазмы крови а,-антитрипсин может инактивировать некоторые протеазы, освобождающиеся в острой фазе воспаления.
Содержание некоторых белков в плазме крови и их функции представлены в таблице 14-2.
Альбумин. Концентрация альбумина в крови составляет 40-50 г/л. В сутки в печени синтезируется около 12 г альбумина, Т1/2 этого белка - примерно 20 дней. Альбумин состоит из 585 аминокислотных остатков, имеет 17 дисульфидных связей и обладает молекулярной массой 69 кД. Молекула альбумина содержит много дикарбоновых аминокислот, поэтому может удерживать в крови катионы Са2+, Cu2+, Zn2+. Около 40% альбумина содержится в крови и остальные 60% в межклеточной жидкости, однако его концентрация в плазме выше, чем в межклеточной жидкости, поскольку объём последней превышает объём плазмы в 4 раза.
Благодаря относительно небольшой молекулярной массе и высокой концентрации альбумин обеспечивает до 80% осмотического давления плазмы. При гипоальбуминемии осмотическое давление плазмы крови снижается. Это приводит к нарушению равновесия в распределении внеклеточной жидкости между сосудистым руслом и межклеточным пространством. Клинически это проявляется как отёк. Относительное снижение объёма плазмы крови сопровождается снижением почечного кровотока, что вызывает стимуляцию системы ренинангиотензинальдрстерон, обеспечивающей восстановление объёма крови (см. раздел 11). Однако при недостатке альбумина, который должен удерживать Na+, другие катионы и воду, вода уходит в межклеточное пространство, усиливая отёки.
Гипоальбуминемия может наблюдаться и в результате снижения синтеза альбуминов при заболеваниях печени (цирроз), при повышении проницаемости капилляров, при потерях белка из-за обширных ожогов или катаболических состояний (тяжёлый сепсис, злокачественные новообразования), при нефротическом синдроме, сопровождающемся альбуминурией, и голодании. Нарушения кровообращения, характеризующиеся замедлением кровотока, приводят к увеличению поступления альбумина в межклеточное пространство и появлению отёков. Быстрое увеличение

237
проницаемости капилляров сопровождается резким уменьшением объёма крови, что приводит к падению АД и клинически проявляется как шок.
Альбумин - важнейший транспортный белок. Он транспортирует свободные жирные кислоты (см. раздел 8), неконъюгированный билирубин (см. раздел 13), Са2+, Сu2+, триптофан, тироксин и трийодтиронин (см. раздел 11). Многие лекарства (аспирин, дикумарол, сульфаниламиды) связываются в крови с альбумином. Этот факт необходимо учитывать при лечении заболеваний, сопровождающихся гипоальбуминемией, так как в этих случаях повышается концентрация свободного лекарства в крови. Кроме того, следует помнить, что некоторые лекарства могут конкурировать за центры связывания в молекуле альбумина с билирубином и между собой.
Транстиретин (преальбумин) называют тирок-синсвязывающим преальбумином. Это белок острой фазы. Транстиретин относят к фракции альбуминов, он имеет тетрамерную молекулу. Он способен присоединять в одном центре связывания ретинолсвязывающий белок, а в другом - до двух молекул тироксина и трийодтиронина.
Таблица 14-2. Содержание и функции некоторых белков плазмы крови
|
|
Концентрация в |
|
|
|
|
Группа |
Белки |
сыворотке крови, |
Функция |
|
|
|
|
|
г/л |
|
|
|
|
|
|
|
|
|
|
|
Альбумины |
Транстиретин |
0,25 |
Транспорт |
тироксина |
и |
|
|
|
|
трийодтиронина |
|
|
|
|
|
|
|
|||
|
Альбумин |
40 |
Поддержание осмотического давления, |
|||
|
|
|
транспорт жирных кислот, билирубина, |
|||
|
|
|
жёлчных |
кислот, |
стероидных |
|
|
|
|
гормонов, лекарств, неорганических |
|||
|
|
|
ионов, резерв аминокислот |
|
|
|
|
|
|
|
|
|
|
α1-Глобулины |
α1 -Антитрипсин |
2,5 |
Ингибитор протеиназ |
|
|
|
|
|
|
|
|
|
|
|
ЛПВП |
0,35 |
Транспорт холестерола |
|
|
|
|
|
|
|
|
||
|
Протромбин |
0,1 |
Фактор II свёртывания крови |
|
||
|
|
|
|
|||
|
Транскортин |
0,03 |
Транспорт кортизола, кортикостерона, |
|||
|
|
|
прогестерона |
|
|
|
|
|
|
|
|
|
|
|
Кислый α1-гликопротеин |
1 |
Транспорт прогестерона |
|
|
|
|
|
|
|
|
|
|
|
Тироксинсвязывающий |
0,02 |
Транспорт |
тироксина |
и |
|
|
глобулин |
|
трийодтиронина |
|
|
|
|
|
|
|
|
|
|
α2-Глобулины |
Церулоплазмин |
0,35 |
Транспорт |
ионов |
|
меди, |
|
|
|
оксидоредуктаза |
|
|
|
|
|
|
|
|
||
|
Антитромбин III |
0,3 |
Ингибитор плазменных протеаз |
|
||
|
|
|
|
|
|
|
|
Гаптоглобин |
1 |
Связывание гемоглобина |
|
|
|
|
|
|
|
|
|
|
|
α2-Макроглобулин |
2,6 |
Ингибитор |
плазменных |
протеиназ, |
|
|
|
|
транспорт цинка |
|
|
|
|
|
|
|
|
|
|
|
Ретинолсвязыва-ющий |
0,04 |
Транспорт ретинола |
|
|
|
|
белок |
|
|
|
|
|
|
|
|
|
|
|
|

|
|
|
|
238 |
|
|
|
|
|
|
|
|
Витамин |
D |
0,4 |
|
Транспорт кальциферола |
|
связывающий белок |
|
|
|
|
|
|
|
|
|
|
β-Глобулины |
ЛПНП |
|
3,5 |
|
Транспорт холестерола |
|
|
|
|
|
|
|
Трансферрин |
3 |
|
Транспорт ионов железа |
|
|
|
|
|
|
|
|
Фибриноген |
3 |
|
Фактор I свёртывания крови |
|
|
|
|
|
|
|
|
Транскобаламин |
25×10-9 |
|
Транспорт витамина B12 |
|
|
|
|
|
|
|
|
Глобулин |
связывающий |
20×10-6 |
|
Транспорт тестостерона и эстрадиола |
|
белок |
|
|
|
|
|
|
|
|
|
|
|
С-реактивный белок |
<0,01 |
|
Активация комплемента |
|
|
|
|
|
|
|
γ-Глобулины |
IgG |
|
12 |
|
Поздние антитела |
|
|
|
|
|
|
|
IgA |
|
3,5 |
|
Антитела, защищающие слизистые |
|
|
|
|
|
оболочки |
|
|
|
|
|
|
|
IgM |
|
1,3 |
|
Ранние антитела |
|
|
|
|
|
|
|
IgD |
|
0,03 |
|
Рецепторы В-лимфоцитов |
|
|
|
|
|
|
|
IgE |
|
<0,01 |
|
Реагин |
|
|
|
|
|
|
Соединение с этими лигандами происходит независимо друг от друга. В транспорте последних транстиретин играет существенно меНbшую роль по сравнению с тироксинсвязывающим глобулином.
α1 - Антитрипсин относят к α1-глобулинам. Он ингибирует ряд протеаз, в том числе фермент эластазу, освобождающийся из нейтрофилов и разрушающий эластин альвеол лёгких. При недостаточности α1-антитрипсина могут возникнуть эмфизема лёгких (см. раздел 15) и гепатит, приводящий к циррозу печени. Существует несколько полиморфных форм α1-антитрипсина, одна из которых является патологической. У людей, гомозиготных по двум дефектным аллелям гена антитрипсина, в печени синтезируется α1-антитрипсин, который образует агрегаты, разрушающие гепатоциты. Это приводит к нарушению секреции такого белка гепатоцитами и к снижению содержания α1-антитрипсина в крови.
Гаптоглобин составляет примерно четверть всех α2-глобулинов. Гаптоглобин при внутрисосудистом гемолизе эритроцитов образует комплекс с гемоглобином, который разрушается в клетках РЭС. Если свободный гемоглобин, имеющий молекулярную массу 65 кД, может фильтроваться через почечные клубочки или агрегировать в них, то комплекс гемоглобин-гаптоглобин имеет слишком большую молекулярную массу (155 кД), чтобы пройти через гломерулы. Следовательно, образование такого комплекса предотвращает потери организмом железа, содержащегося в гемоглобине. Определение содержания гаптоглобина имеет диагностическое значение, например, снижение концентрации гаптоглобина в крови наблюдают при гемолитической анемии. Это объясняют тем, что при Т1/2 гаптоглобина, составляющем 5 дней, и Т1/2 комплекса гемоглобин - гаптоглобин (около 90 мин) увеличение поступления свободного гемоглобина в кровь при гемолизе эритроцитов вызовет резкое снижение содержания свободного гаптоглобина в крови.
Гаптоглобин относят к белкам острой фазы, его содержание в крови повышается при острых воспалительных заболеваниях.
Информация о некоторых других белках плазмы крови, представленных в табл. 14-2, имеется в соответствующих разделах учебника.
239
Энзимодиагностика
Энзимодиагностика заключается в постановке диагноза заболевания (или синдрома) на основе определения активности ферментов в биологических жидкостях человека. Принципы энзимодиагностики основаны на следующих позициях:
при повреждении клеток в крови или других биологических жидкостях (например, в моче) увеличивается концентрация внутриклеточных ферментов повреждённых клеток;
количество высвобождаемого фермента достаточно для его обнаружения;
активность ферментов в биологических жидкостях, обнаруживаемых при повреждении клеток, стабильна в течение достаточно длительного времени И отличается от нормальных значений;
ряд ферментов имеет преимущественную или абсолютную локализацию в определённых органах (органоспецифичность);
существуют различия во внутриклеточной локализации ряда ферментов.
1. |
Причины, |
приводящие |
к |
увеличению |
количества |
ферментов в крови |
|
|
|
|
|
Ферменты плазмы крови можно разделить на 2 группы. Первая, относительно небольшая группа ферментов активно секретируется в плазму крови определёнными органами. Например, печень синтезирует неактивные предшественники ферментов свёртывающей системы крови. Ко второй относят большую группу ферментов, высвобождающихся из клеток во время их нормального функционирования. Обычно эти ферменты выполняют свою функцию внутри клетки и не имеют физиологического значения в плазме крови. У здорового человека активность этих ферментов в плазме низкая и достаточно постоянная, так как постоянно соотношение скоростей высвобождения их из клеток и скоростей разрушения.
При многих заболеваниях происходит повреждение клеток, и их содержимое, в том числе и ферменты, высвобождаются в кровь. К причинам, вызывающим высвобождение внутриклеточного содержимого в кровь, относят нарушение проницаемости мембраны клеток (при воспалительных процессах) или нарушение целостности клеток (при некрозе). Определение в крови активности ряда ферментов хорошо налажено в биохимических лабораториях, что используют для диагностики заболеваний сердца, печени, скелетной мускулатуры и других тканей. Уровень активности ферментов в плазме коррелирует со степенью повреждения клеток.
Для энзимодиагностики имеют большое значение знания о субклеточной локализации ферментов. Так, появление в плазме крови ферментов, имеющих только цитозольную локализацию, свидетельствует о воспалительном процессе; при обнаружении митохондриальных или ядерных ферментов можно говорить о более глубоких повреждениях клетки, например о некрозе.
Однако повышение концентрации ферментов не всегда связано с повреждением тканей. При избыточной клеточной пролиферации, например при онкопролиферативных процессах, при повышенной скорости синтеза некоторых ферментов в клетках или при нарушенном клиренсе (способности вьпюдиться почками) наблюдают повышение концентрации в крови определённых ферментов. Врачам следует учитывать, что нормальные значения активности ферментов в крови детей и беременных женщин отличаются от показателей, характерных для взрослых здоровых людей.
2. Изоферменты
Ферменты, катализирующие одну и ту же химическую реакцию, но отличающиеся по первичной структуре белка, называют изофермен-тами, или изоэнзимами. Они катализируют один и тот же тип реакции с принципиально одинаковым механизмом, но отличаются друг от друга кинетическими параметрами, условиями активации, особенностями связи апофермента и кофермента.

240
Природа появления изоферментов разнообразна, но чаще всего обусловлена различиями в структуре генов, кодирующих эти изоферменты. Следовательно, изоферменты различаются по первичной структуре белковой молекулы и, соответственно, по физико-химическим свойствам. На различиях в физико-химических свойствах основаны методы определения изоферментов.
По своей структуре изоферменты в основном являются олигомерными белками. Причём та или иная ткань преимущественно синтезирует определённые виды протомеров. В результате определённой комбинации этих протомеров формируются ферменты с различной структурой - изомерные формы. Обнаружение определённых изоферментных форм ферментов позволяет использовать их для диагностики заболеваний.
Изоформы лактатдегидрогеназы. Фермент лак-татдегидрогеназа (ЛДГ) катализирует обратимую реакцию окисления лактата (молочной кислоты) до пирувата (пировиноградной кислоты) (см. раздел 7).
Лактатдегидрогеназа - олигомерный белок с молекулярной массой 134 000 Д, состоящий из 4 субъединиц 2 типов: М (от англ, muscle - мышца) и Н (от англ, heart - сердце). Комбинация этих субъединиц лежит в основе формирования 5 изоформ лактатдегидрогеназы (рис. 2-35, А). ЛДГ1 и ЛДГ2 наиболее активны в сердечной мышце и почках, ЛДГ4 и ЛДГ5 - в скелетных мышцах и печени.
Востальных тканях имеются различные формы этого фермента.
Изоформы ЛДГ отличаются электрофоретической подвижностью, что позволяет устанавливать тканевую принадлежность изоформ ЛДГ (рис. 2-35, Б).
Рис. 2-35. Изоформы лактатдегидрогеназы. А - строение различных изоформ ЛДГ; Б -
распределение на электрофореграмме и относительные количества изоформ ЛДГ в различных органах; В - содержание изоформ ЛДГ в плазме крови в норме и при патологии (электрофореграммы - слева и фотометрическое сканирование - справа).