

Липостат.
Липостат (массостат) —условное название системы, контролирующей постоянство веса тела. Как и для других балансовых констант организма —температуры, осмолярности тканевой жидкости, кровяного давления центральным контролирующим звеном в системе, регулирующей массу тела, служит гипоталамус. Липостатический гомеостаз обеспечивается путем прямых и обратных сигнальных взаимодействий между гипоталамусом и жировой тканью (с её гормонами), а также гипоталамусом и ЖКТ (с его энтериновой гормональной системой). (См. рис. 2.3.)
4.1. Основные звенья и механизмы функционирования липостата.
 Идею
о существовании «пищевого центра»,
как автоматического регулятора
потребления пищи, впервые высказал
И.П. Павлов (1911).Он
считал, что в коре больших полушарий
и нижележащих отделах ЦНС имеется
функциональное объединение клеток,
чувствительных к степени наполнения
желудка и химическим сигналам, и
обеспечивающих контроль поступления
пищи. Ранние представления об обратной
афферентации в системе контроля пищевого
поведения связывали сигнал сытости
со степенью растяжения желудка и
обратной афферентацией от механических
процессов, сопутствующих обработке
пищи во рту и ее пассажу через глотку и
пищевод(механическая теориярегуляции аппетита). Установлено,
что эти факторы действительно вносят
вклад в краткосрочные механизмы
регуляции потребления пищи.
Идею
о существовании «пищевого центра»,
как автоматического регулятора
потребления пищи, впервые высказал
И.П. Павлов (1911).Он
считал, что в коре больших полушарий
и нижележащих отделах ЦНС имеется
функциональное объединение клеток,
чувствительных к степени наполнения
желудка и химическим сигналам, и
обеспечивающих контроль поступления
пищи. Ранние представления об обратной
афферентации в системе контроля пищевого
поведения связывали сигнал сытости
со степенью растяжения желудка и
обратной афферентацией от механических
процессов, сопутствующих обработке
пищи во рту и ее пассажу через глотку и
пищевод(механическая теориярегуляции аппетита). Установлено,
что эти факторы действительно вносят
вклад в краткосрочные механизмы
регуляции потребления пищи.
Рисунок 2.3. Схема
регуляции липостата. [4;2000]
Кишечные гормоны энтериновой системы выделяются в ответ на прием пищи и подавляют чувство аппетита. Особенно активны гормоны двенадцатиперстной кишки. Одна из гормонально активных фракций экстрактов двенадцатиперстной кишки, содержащая сигнальные пептиды, подавляющие активность центра голода, была названа арэнтерином. Ученые показали, что аппетит-регулирующий энтериновыи гормон имеет белковую природу и молекулярную массу около 100кД
Сильным ингибитором чувства голода, аппетита и пищевой активности служит дуоденальный гормон холецистокинин Потребление воды холецистокинин не затрагивает.Данный гормон обладает центральным тормозным действием на вентолатеральный гипоталамус, а также действует, опосредованно через рецепторы и афферентные волокна абдоминальной вагус-ной системы
Менее активными ингибиторами центра голода являются бомбезин, соматостатин, сатиетин, нейротензин, кортиколиберин, тиролиберин, вазоактивный интестинальный полипептид и инсулин, имеющие рецепторы в центре насыщения. Дериват тиролиберина —гистидилпролин дикетопиперазин —понижает аппетит, вызывает истощение у крыс и обнаружен в крови больных психогенной анорексией. Эндорфины и энкефалины, а также соматолиберин - наоборот, стимулируют аппетит. Все эти регуляторы вырабатываются как в энтериновой системе, так и в ЦНС.
Энтериновые гормоны, понижающие возбуждение центра голода, выделяются в ответ на попадание пищи или даже просто —кислого желудочного сока —в двенадцатиперстную кишку.
Аппетит и насыщение подвержены действию ряда нейротрансмиттеров. Норадреналин, известный как липолитический активатор, тормозит аппетит; вызывают чувство насыщения и другие -адреномиметики, что используется при применении амфетаминов в качестве анорексигенов. Менее активен в качестве анорсксигена серотонин.
В процессах липостата участвует также и жировая ткань.
Жировая тканьпринадлежит к соединительным тканям со специальными свойствами, вместе с ретикулярной и пигментной тканями, а также слизистой тканью вартонова студня и стекловидного тела.Клетки жировой ткани —адипоциты —формируются из преадипоцитов, которые, в свою очередь, являются потомками фибробластов. Дифференцировка преадипоцитов в новые адипоциты интенсивно идет в последний триместр внутриутробной жизни и продолжается до 3-го месяца постнатального периода. При дифференцировке пре адипоциты экспрессируют маркерный фермент липопротеиновую липазу (ЛПЛ), утрачивают отростки, синтезируют коллагены I и XIтипов, некоторые белки цитоскелета и обретают несколько жировых капель, которые затем в белом жире сливаются в одну, а буром —остаются раздельными. При голодании, теряя жир, адипоциты вновь приобретают фибробластоподобный вид, а при откармливании процесс их дифференцировки как бы повторяется. Дифференцировке адипоцитов способствуют соматотропный гормон и инсулиноподобный фактор роста .Тиреоидные гормоны сдерживают этот процесс.
Адипоциты динамично реагируют на изменения эндокринно-метаболической ситуации, так как имеют обширный набор поверхностных нейромедиаторных и гормональных рецепторов. В этих клетках неприрывно идут процессы липогенеза и липолиза. [4;2000]
Липогенез —процесс синтеза жирных кислот, интенсивно протекающий в печени и жировой ткани. Следует отметить, что биосинтез липидов осуществляется в основном в цитоплазме клетки и лишь незначительная их часть, как считают некоторые авторы, образуется в митохондриях.
Образовавшийся в митохондриях ацетил-СоА способен проникать через их мембрану в цитоплазму только после взаимодействия с оксалоацетатом и образования цитрата, который достигает цитоплазмы, где вновь превращается в ацетил-СоА и оксалоацетат. Таким образом, оксалоацетат выполняет функцию катализатора, способствуя переносу ацетильной группы из интрамитохондриального пространства в цитозоль, где при участии ацетил-СоА-карбоксилазы происходит карбоксилирование ацетил-СоА с образованием малонил-СоА. Последний, взаимодействуя с комплексом жирные кислоты —белок, при участии АТФ и СоА превращается в комплекс жирнокислотный ацетил-СоА —необходимую стадию образования триглицеридов.
Карбоксилирование ацетил-СоА является важным регулятором синтеза жирных кислот, и на всех последующих этапах липогенеза участвует либо малонил-СоА, либо ацетил-СоА. Так, под влиянием синтетазы жирных кислот и при использовании одной молекулы ацетил-КоА и молекул малонил-СоА образуется пальмитил-СоА, способный к разнообразным превращениям. При потере одной молекулы СО, и СоА образуется пальмитиновая кислота, молекула которой содержит 16атомов углерода. Путем удлинения цепи в митохондриях или в плазматическом ретикулуме до 18атомов углерода пальмитиновая кислота может превращаться в стеариновую, а после денатурации она же преобразуется в пальмитоолеиновую и олеиновую кислоты.
При эстерификации жирных кислот с глицерином образуются триглицериды, причем в реакции участвует не свободный глицерин, а его производное —глицерол-3-фосфат, образующийся в печени из глицерина при участии глицеролкиназы. Эстерификация жирных кислот в жировой ткани может происходить лишь при достаточном поступлении дигидроксиацетонфосфата, который образуется в процессе гликолиза и конвертируется в глицерол-3-фосфат при участии глицерофосфатдегидрогеназы.
В жировой ткани также осуществляется и из глюкозы. Инсулин служит мощным стимулятором этого процесса, увеличивая захват жировой тканью глюкозы, активность фосфофруктокиназы, пропускную способность фосфотриозного и пентозного пути метаболизма. Глюкогон влияет на эти процессы противоположным образом. Глюкокортикоиды увеличивают и липогенез, и липолиз, причём суммарный эффект их действия различен в адипоцитах разной локализации, в силу пермиссивного действия катехоламинов на жировые клетки, имеющие неодинаковый набор адренорецепторов. При гиперкортицизме подкожный жир откладывается на лице,шее и в верхней части туловища. На конечностях количество жира уменьшается. [4;2000,2;2000]
Липолиз —процесс гидролиза липидов в жировой ткани с образованием неэстерифицированных (свободных) жирных кислот и глицерола (глицерина) —катализируется двумя группами внутриклеточных ферментов: гормонально-зависимой липазой и другими триацилглицероловыми липазами (диацилглицероловая и моноацилглицероловая липазы)(рис.2.4.)
Гормональнозависимая липаза, стимулируемая цАМФ, осуществляет отщепление от триацилглицерола первой жирной кислоты и высвобождает диацил глицерин (или диацилглицерол). Гормонально-зависимая липаза лимитирует скорость липолиза, а образование цАМФ под влиянием аденилатциклазы находится в свою очередь под контролем различных гормонов.
Гормональнозависимая липаза по своим свойствам отличается от других липаз: ее активность регулируется реверсивным (обратным) фосфорилированием, и эта липаза обладает одинаковой каталитической активностью как в отношении триацилглицерола, так и в отношении длинноцепочечных эфиров холестерина. Другие триацилглицероловые липазы не обладают холестерилэстеразной активностью. Гидролиз триацилглицеролов происходит более медленно, чем диацилглицерола. Поэтому многие специалисты полагают, что лимитирующим скорость этапом в липолизе жира является гидролиз триацилглицерола.
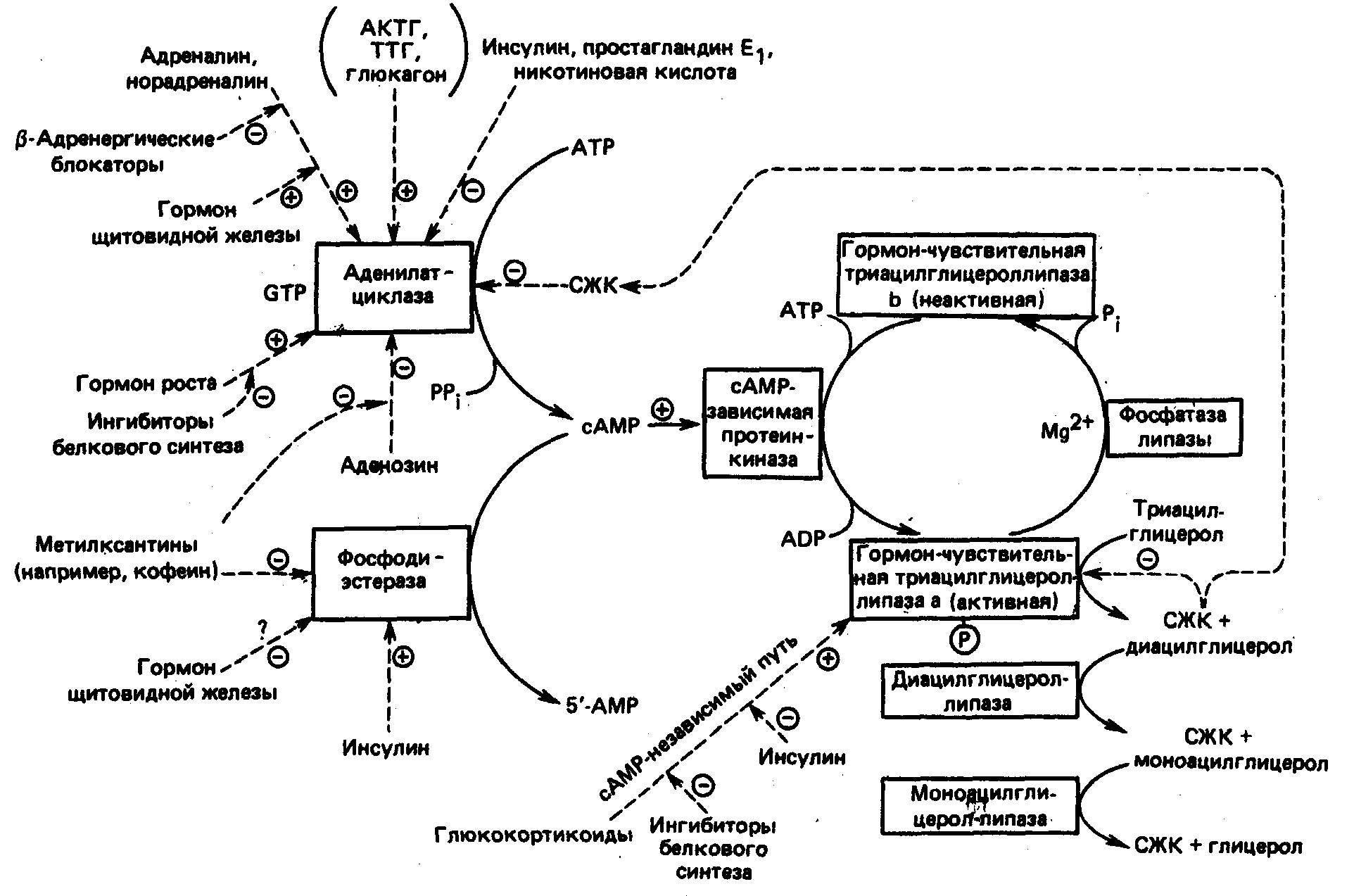
Рисунок 2.4. Регуляция липолиза в жировой ткани. ТТГ-тиреотропный гормон: СЖК - свободные жирные кислоты. [ 8;1993]
Мембрана адипоцитов содержит рецепторы, взаимодействующие с гормонами, обладающими липолитическими свойствами (катехоламины, глюкокортикоиды, СТГ, тироксин), и рецепторы к инсулину. Необходимо отметить, что мембрана адипоцитов отличается от других клеточных мембран тем, что в ее состав входят уникальные белки, названные перилипинами. Точная роль этих белков пока не известна, но предполагают, что они принимают участие в "упаковке" нейтральных липидов или в их гидролизе, а может быть, активно участвуют в регуляции этих двух процессов. Перилипины, помимо адипоцитов, выявляются в мембране клеток, секретирующих стероидные гормоны. В других клетках организма они отсутствуют. Структура перилипинов включает около 100аминокислотных остатков, и их последовательность не имеет гомологии с известными другими белками.
В результате действия липолитических гормонов —катехоламинов, СТГ, глюкокортикоидов и тироксина (глюкагон у человека практически не влияет на липолиз) повышается активность аденилатциклазы, увеличивается образование цАМФ, активируются гормонально-зависимая и другие липазы, участвуя, таким образом, в липолизе жира. Взаимодействие инсулина с соответствующими рецепторами, наоборот, приводит к угнетению аденилатциклазы, снижению концентрации цАМФ и торможению липолиза. Липолиз увеличивается во время голодания, при продолжительной работе, охлаждении, стрессе. Липолитическое действие катехоламинов (адреналин, норадреналин) и глюкагона осуществляется путем активации аденилатциклазы. С физиологической точки зрения, роль норадреналина в процессе липолиза представляется более важной, чем адреналина. Он образуется в адренергических нервных окончаниях в жировой ткани и обеспечивает мобилизацию жирных кислот. Катехоламины оказывают свое липолитическое действие через-адренорецепторы, повышая активность аденилатциклазы и уровень цАМФ, тогда как активирование-адренергических рецепторов под влиянием катехоламинов сопровождается ингибированием липолиза. Особая роль в процессах липолиза под влиянием катехоламинов отводится "нетипичным"-адренорецепторам, которые посредством ГТФ-связывающихG-белковкомплексируются с аденилатциклазой. Последующее повышение уровня цАМФ стимулирует гормонально-чувствительную липазу и индуцирует высвобождение неэстерифицированных жирных кислот, которые подвергаются затем окислению.
Гормон роста оказывает мощное липолитическое действие, которое отличается от действия катехоламинов. Он вызывает увеличение концентрации свободных жирных кислот в плазме через 2—3ч. Это действие, отмечающееся при введении даже небольших доз СТГ, по-видимому, связано с торможением процесса реэстерификации свободных жирных кислот. Тем не менее СТГ оказывает свое определенное модулирующее влияние и на активность аденилатциклазы. Однако основное липолитическое действие СТГ, как и тироксина, не зависит от цАМФ. [2;2000]
Другие гипофизарные гормоны (АКТГ, ТТГ, меланоцитостимулирующий гормон) также оказывают липолитическое действие, хотя и менее выраженное, чем у СТГ. Большинство авторов считают, что оно осуществляется путем активации аденилатциклазы и повышения скорости образования цАМФ.
Тироидные гормоны и кортикостероиды оказывают пермиссивное (разрешающее) действие на липолиз, поскольку липолитический и калоригенный эффекты катехоламинов не проявляются в отсутствие кортикостерондов и тироидных гормонов. Глюкокортикоиды значительно потенцируют липолитическое действие адреналина. Имеются данные о том, что ТТГ, АКТГ, гормоны щитовидной железы и коры надпочечников индуцируют синтез аденилатциклазы и, следовательно, принимают непосредственное участие в стимуляции липолиза. Кроме того, тироидные гормоны влияют на активность ацил-КоА-синтетазы, локализующейся в митохондриях, и цитоплазматического фермента глицеролфосфатдегидрогеназы. Эти ферменты участвуют в регуляции скорости липолиза и липогенеза. При гипопитуитаризме, не затрагивающем функции гипоталамуса, возможно развитие вторичного ожирения. Жирные кислоты и глицерин освобождаются в плазму. Первые связываются альбумином для транспорта в гепатоииты. Но около 40%глицерина реэтерифицируется с жирными кислотами, поставляемыми ЛПЛ. Интересно, что у лице наклонностью к первичному ожирению значительно большая доля глицерина подвергается реэтерификации в ТГ, а меньшая —освобождается. В связи с этим, существовала даже гипотеза, что нормальная активность липостата и его правильная с антропометрической точки зрения установка, относительно массы тела, достигаются лишь при адекватном уровне глицерина в крови. Делались, в русле этих представлений, попытки глицеринотерапии первичного ожирения. Роль глицерина как ведущего сигнала обратной афферентации в липостате, не была подтверждена,
Инсулин обладает характерным антилиполитическим свойством, и при сахарном диабете вследствие увеличения липолиза повышается концентрация свободных жирных кислот в плазме, снижению которой способствует инсулинотерапия. Инсулин угнетает липолиз посредством снижения активности гормонально-зависимой липазы и повышает, таким образом, поглощение глюкозы и уровень глицерол-3-фосфата. Показано, что повышение содержания инсулина в плазме крови всего на 10—14мкЕД/мл выше базального его уровня приводит к резкому снижению концентрации свободных жирных крови. [2;2000]
Также важную роль в процессе липостата играет пептидный гормон, вырабатываемый адипоцитами - лептин.