

30. Открытие групп крови системы ав0 к. Ландштейнером. Характеристика групп крови системы ав0. Резус-система и ее характеристика.
В начале XX века Ландштейнер обнаружил, что в плазме (сыворотке) крови кролика могут вырабатываться факторы, которые вызывают склеивание эритроцитов (агглютинацию) крови человека. В 1901 году К. Ландштейнер впервые открыл группы АВ0, 1927 году совместно с Левиным он открыл факторы N, M, P, в 1937-1940гг. совместно с А. Винером открыл резус-фактор.
Система АВ0. Кровь как ткань обладает антигенными свойствами. АГ-АТ. Мембрана эритроцитов содержит определенные антигены – агглютиногены А, В; в плазме – антитела – агглютинины α, β.
|
Группа |
Агглютиногены (АГ) |
Агглютинины (АТ) |
|
I 0 |
- |
α, β |
|
II A |
A |
Β |
|
III В |
В |
Α |
|
IV AB |
A, В |
- |
Содержание АГ и АТ передается по наследству.
IA, IB, i
I 0 - ii
II A - IAIA, IAi
III В – IBIB, IBi
IV AB - IAIB
Это основная серологическая система, определяющая совместимость или несовместимость крови при ее переливании. В нее входят два генетически детерминированных агглютиногена А и В и два агглютинина – α и β.
Существуют варианты антигена А – А1, А2, А3, А4 и т.д., из которых А1 – сильный антиген, А2, А3 и др. – более слабые. В связи с этим группа Аβ (II) неоднородна – у 88% содержится сильный антиген А1, а у 12% - А2. Это имеет принципиальное значение при определении группы крови: люди, имеющие агглютиноген А2, могут быть приняты за человека с группой 0 α β (I).
Существуют варианты антигена В – В1, В2, В3 и т.д., но все эти антигены по своей антигенной активности равны между собой, поэтому при определении группы крови Bα (III), как правило, ошибок не бывает. У людей с группой крови АВ0 (IV) также могут встречаться разные агглютиногены А, поэтому часть этих людей может быть определена как человек с группой Вα (III) при условии, что в эритроцитах у них содержится слабый в антигенном отношении агглютиноген А2.
Антитела α- и β-агглютинины относятся к классу иммуноглобулинов IgM, они проходят через плаценту, являются нормальными (естественными), полными, холодовыми и термолабильными, т.е. разрушаются при 70°С.
Определение группы крови по системе АВ0 проводят различными способами, в том числе по стандартным изогемагглютинирующим сывороткам I, II, III групп: используются два ряда стандартных сывороток. В каплю сыворотки вносятся эритроциты исследуемого в соотношении 10:1. Реакция проводится при комнатной температуре. Существует современный способ, основанный на использовании моноклониальных антител – цоликлон анти-А и цоликлон анти-В. Этот метод позволяет избежать ошибок, возможных из-за наличия слабых антигенов типа А2. Для надежного определения групповой принадлежности в сомнительных случаях, например, при подозрении на наличие А2, используется перекрестный метод – к стандартным эритроцитам I, II и III групп добавляется исследуемая сыворотка. Во всех методиках критерием оценки является появление в соответствующих случаях агглютинации эритроцитов.
|
Ген |
Антигены на мембране эритроцита |
Антитела (в плазме крови) |
Группа крови системы АВ0 |
|
Н(0) |
Н |
Анти-А+Анти-А2+Анти-В |
I (0) |
|
A1 |
A1 |
Анти-В |
II (A1) |
|
A2 |
A2+H |
Анти-А1 у 1% обследуемых+Анти-В |
II (A2) |
|
В |
В |
В |
III (B) |
|
A1, B |
A1+B |
Анти-А+Анти-А1 |
IV (A1, B) |
|
A2, B |
A2+B |
Отсутствует Анти-А1 у 25% обследуемых |
IV (A2, B) |
Система резус.
Открыта в результате иммунизации кроликов кровью обезьян – макак-резусов (Ландштейнер, Винер, 1937-1940). В настоящее время выявлено много антигенов этой системы, но их иммуногенная сила разная. Существуют две основных номенклатуры обозначения антигенов этой системы: по Ландштейнеру и Винеру и по Фишеру Р. И Райсу Р. Современная номенклатура – это совмещение двух номенклатур.
Антигены. Современный вариант: Rh0 (D): rhI (C): rhII (E): Hr0 (d): hrI (c): hrII (e).
Наиболее активным в антигенном отношении является антиген D, в меньшей степени – С и Е, а тем более d, c, e. Реципиент имеет резус-положительную кровь, если его эритроциты обязательно содержат антиген D. Антиген D выявляется у 86% людей, С – у 70,8%, Е – у 31,0%, d – y 99%, с – у 84%, е – у 86%. Учитывая, что антиген D определяет принадлежность людей к группе резус-положительных, таких людей среди европейцев много – 86%, у представителей монгольской расы – 100%.
Антиген D является основной причиной сенсибилизации (иммунизации) во время беременности и гемолитической болезни новорожденных, он легко проникает через плаценту.
В настоящее время известны и другие факторы резус-системы. Из них особый интерес представляет вариант фактора D, который обозначается Du. Он не всегда определяется в эритроцитах, но в ответ на его введение у резус-отрицательного человека вырабатывается анти-D. Поэтому у резус-отрицательного человека необходимо определить и отсутствие антигена Du.
В эритроците антигены в системы резус находятся в виде группы антигенов. Наиболее частые комбинации такие: CDE – 16%, CDe – 53%, cDE – 15%, cde – 12%. У аборигенов Австралии в эритроцитах не выявлен ни один представитель системы резус. Такой вариант называют резус-нуль.
Для определения резус-принадлежности, т.е. выявления антигенов системы резус в эритроцитах используют стандартные сыворотки (реагенты) антирезус, различные по специфичности, т.е. содержащие антитела к разным антигенам этой системы. Для определения антигена D чаще всего применяют сыворотку антирезус с добавлением 10% раствора желатина или используют стандартный реагент антирезус, приготовленный заранее с 33% раствором полиглюкина.
31. Функциональная система поддержания оптимального для метаболизма pO2/pCO2. Транспорт кислорода. Кривая диссоциации оксигемоглобина. Транспорт углекислого газа, участие эритроцитов в транспорте CO2.
Система, которая поддерживает оптимальное соотношение O2/CO2.
ЦНС (гипоталамус, кора дыхательного центра, продолговатый мозг)→внешнее дыхание; работа сердечно-сосудистой системы, количество эритроцитов, гемоглобин, регуляция рН, уровень метаболических процессов, функция почек (механизм регуляции – гормоны)→ O2/CO2→хеморецепторы рН.
Транспорт кислорода.
Кислород транспортируется гемоглобином эритроцитов. 1г-1,34 мл O2, Hb=150 г/л, 134+67=200 О2.
Кислород диффундирует из альвеол в кровь легочных капилляров. В небольшом количестве кислород транспортируется кровью в физически растворенном состоянии. Поскольку растворимость кислорода в жидкости при 37°С составляет 0,225 мл · л-1 · кПа-1 (0,03 мл/л/мм рт. с.), то при нормальном напряжении в альвеолярном воздухе, т.е. кислород 13,3 кПа или 100 мм рт. ст., каждые 100 мл плазмы крови могут переносить 0,3 мл кислорода, что недостаточно для жизнедеятельности организма.
Кривая диссоциации оксигемоглобина.
Кислорода в плазме растворяется очень мало (0,02-0,03). Большая часть транспортируется в крови в соединении с гемоглобином - в форме оксигемоглобина. Гемоглобин имеет разное сродство к кислороду. Это сродство зависит от парциального давления. Эта зависимость выражается кривой диссоциации оксигемоглобина. Молекула кислорода соединяется прочно и обратимо с гемом гемоглобина. Эта связь носит нелинейный характер и выражается кривой диссоциации оксигемоглобина, имеющей S-образную форму. Кривая характеризует процент сатурации или насыщения гемоглобина кислородом. Процент оксигемоглобина в артериальной крови в среднем равен 95-97%. Ацидоз – прямая сдвигается вправо (гиперкапния, уменьшение рН)
Транспорт CO2.
Осуществляется в 3-х основных формах, но самая главная – 1. соль угольной кислоты. CO2+H2O=H2CO3+K+=KHCO3+H+.
2. CO2+Hb=HbCO2 – транспортируется порядка 30% CO2$ карбогемоглобин.
3. Небольшое кол-во CO2 может раствориться в плазме (несколько мм – 1л плазмы)
Эритроциты и гемоглобин играют важную роль в транспорте СО2. Попадание СО2 в эритроцит увеличивает подачу кислорода в ткани.
Гипокапния – мало СО2 в крови; гиперкапния – создает благоприятные условия для отдачи кислорода в ткани, много СО2.
32. Система внешнего дыхания, дыхательные пути и их характеристика. Дыхательные мышцы и биомеханика дыхательных движений. Дыхательные объемы и методы их измерения. Частота дыхания, минутный объем дыхания. Альвеолярная вентиляция. Газовый состав атмосферного, выдыхаемого и альвеолярного воздуха. Газообмен в легких.
Дыхание – физиологическая система, представленная совокупностью процессов, которые обеспечивают поступление к клеткам организма кислорода и удаление из организма углекислого газа. Основная функция дыхания - поддержание газового гомеостаза внутренней среды организма в соответствии с уровнем его метаболизма. Физиологические процессы системы дыхания: 1) внешнее дыхание; 2) транспорт О2 от легких к тканям и СО2 в противоположном направлении; 30 газообмен между кровью и тканями. В свою очередь процесс внешнего дыхания состоит из двух стадий: 1) газообмен О2 и СО2 между атмосферным воздухом и альвеолами; 2) газообмен между альвеолами и кровью легочных капилляров.
Система внешнего дыхания.
- система работает ритмично, автономно; средняя частота дыхания 15-16/мин;
- в состоянии покоя в норме вдыхаем 500 мл (0,5л), но не весь воздух будет попадать в легкие (150 мл не доходит, остается в воздухоносных путях – носовой полости, гортани). 150 мл –анатомически мертвое пространство.
Воздух в легких: 16-17% О2; 3-4% СО2.
Функцией процесса внешнего дыхания является газообмен О2 и СО2 между атмосферным и альвеолярным воздухом, который осуществляется с помощью билмеханизма вдоха (увеличивает объем легких) и выдоха (уменьшает объем легких), приводимого в действие сокращением дыхательных мышц. Дыхательные мышцы единственные, которые контролируются как автономно, так и произвольно. Функции дыхательных мышц весьма разнообразны: дыхание, речь, поза.
Дыхательные пути (воздухоносные пути) - это часть аппарата внешнего дыхания, совокупность анатомических структур, представляющих собой дыхательные трубки, по которым смесь дыхательных газов активно транспортируется из среды организма к паренхиме лёгких и обратно - от паренхимы лёгких в среду. Так дыхательные пути участвуют в исполнении функции вентиляции лёгких с целью осуществления внешнего дыхания.
Дыхательные пути разделяют на два отдела: верхние воздухоносные (дыхательные) пути и нижние воздухоносные (дыхательные) пути. Верхние дыхательные пути включают полость носа, носовую часть глотки и ротовую часть глотки. Нижние дыхательные пути включают гортань, трахею и бронхиальное дерево. Бронхиальное дерево представляет собой все внелёгочные и внутрилёгочные ветвления бронхов до концевых бронхиол. Бронхи и бронхиолы подводят и отводят дыхательные газовые смеси к паренхиме лёгких и от нее к верхним дыхательным путям. Паренхима лёгких это часть аппарата внешнего дыхания, состоящая из лёгочных ацинусов. Легочный ацинус начинается концевой (терминальной) бронхиолой, которая ветвится на дыхательные бронхиолы. Дыхательные бронхиолы ветвятся альвеолярными ходами. Альвеолярные ходы завершаются альвеолярными мешочками. Терминальные и дыхательные бронхиолы, а также альвеолярные ходы составляют альвеолярное дерево. Стенки всех элементов альвеолярного дерева составлены альвеолами.
Дыхательные пути и паренхима лёгких являются вероятностной структурой. Как и большинство живых структур, обладают свойством масштабной инвариантности.
В паренхиме лёгких, которую к дыхательным путям не относят, осуществляется циклический процесс внешнего дыхания, частью которого является - диффузионный обмен газами.
Пространство внутри дыхательных путей, объём дыхательных путей, нередко называют анатомическим мертвым пространством, вредным пространством в связи с тем, что в нем не происходит диффузионного обмена газами.
Дыхательные пути выполняют важные функции. Они обеспечивают очищение, увлажнение и согревание вдыхаемой смеси газов (вдыхаемого воздуха).
Биомеханизма вдоха. Во время вдоха объем грудной полости увеличивается в результате сокращения инспираторных мышц: диафрагмы, наружных межреберных и межхрящевых мышц. При сокращении диафрагмальной мышцы купол диафрагмы опускается и одновременно поднимаются и отходят в стороны нижние три ребра, к которым прикреплены ее мышечные волокна. Объем грудной полости увеличивается преимущественно по вертикали. При сокращении наружных межреберных мышц ребра поднимаются, одновременно грудина смещается кпереди.
Биомеханизм выдоха. В первую половину экспирации происходит пассивное управляемое расслабление диафрагмы, одновременно эластическая тяга легких, тяжесть грудной клетки уменьшают объем грудной полости. Экспираторные мышцы (внутренние межреберные и мышцы живота) при спокойном дыхании малоактивны. Однако при вентиляции свыше 40 л/мин, а также при быстрых экспираторных усилиях – глубокое дыхание, кашель, чихание – внутренние межреберные мышцы и мышцы живота активно уменьшают объем грудной клетки. При этом ребра опускаются вниз, так как оказывается больше момент силы, действующий на верхнее ребро. Сокращение мышц живота увеличивает давление в брюшной полости, что также способствует смещению купола диафрагмы вверх.
Дыхательные объемы. Это объемы, которые имеют легкие на вдохе и выдохе при разной глубине дыхательных движений. Дыхательный объем (ДО=500 мл) – объем, который вдыхается или выдыхается при спокойном дыхании. Резервный объем вдоха (РОВд=3000 мл) – максимальное количество воздуха, которое можно вдохнуть после спокойного вдоха. Резервный объем выдоха (РОВ=1100 мл) – максимальное количество воздуха, которое можно выдохнуть после спокойного вдоха. Остаточный объем (ООЛ=1200 мл) – количество воздуха, которое остается в легких после максимального выдоха.
Емкость легких:
1) общая емкость легких (ОЕЛ) – объем воздуха, находящегося в легких после максимального вдоха – все 4 объема (ДО+РОВд+РОВ+OOЛ)=ОЕЛ;
2) жизненная емкость легких (ЖЕЛ) – это объем воздуха, выдохнутого из легких после максимального вдоха при глубоком (максимальном) выдохе. ЖЕЛ=ДО+РОВд+РОВ или ЖЕЛ=ОЕЛ-ООЛ;
3) емкость вдоха (ЕВд) – это максимальный объем воздуха, который можно вдохнуть после спокойного выдоха. ЕВд=ДО+РОВд;
4) функциональная остаточная емкость легких (ФОЕ) – это количество воздуха, остающегося в легких после спокойного выдоха. ФОЕ=РОВ+ООЛ.
Обычно, если выразить эти емкость и объемы в % от ОЕЛ, то ЖЕЛ=70-80% ОЕЛ, ФОЕ – около 50%, ООЛ – около 30%.
Частота — это количество дыхательных актов в минуту. У взрослого человека эта величина обычно составляет 12—15, хотя она может изменяться в широких пределах. У новорожденных частота дыхания во время сна достигает 50—60 в минуту, к годовалому возрасту снижается до 40—50, затем по мере роста происходит постепенное снижение этого показателя. Так, у детей младшего школьного возраста частота дыхания составляет обычно около 25 циклов в минуту, а у подростков — 18—20.
Минутный объем дыхания. Количественным показателем вентиляции легких служит минутный объем дыхания, определяемый как произведение дыхательного объема на число дыханий в минуту. У человека в покое минутный объем дыхания составляет 6-10 л/мин, при работе – от 30 до 100 л/мин. Частота дыхательных движений в покое 12-16 в мин.
Альвеолярная вентиляция. Это часть легочной вентиляции, которая достигает альвеол. Основная функция альвеолярной вентиляции поддерживать необходимую для газообмена концентрацию О2 и СО2 в воздухе альвеол.
Газовый состав.
|
Газы |
Атмосферный воздух (мм рт. ст., %) |
Альвеолярный воздух (мм рт. ст., %) |
Выдыхаемый воздух (мм рт. ст., %) |
|
N2 |
597,0 (78,62%) |
569,0 (74,9) |
566 (74,5) |
|
O2 |
159 (20,84) |
104 (13,6) |
120 (15,7) |
|
CO2 |
0,3 (0,04) |
40 (5,3) |
27 (3,6) |
|
H2O |
3,7 (0,5) |
47 (6,2) |
47 (6,2) |
Газообмен в легких.
В процессе легочного газообмена количество кислорода, поступающего в альвеолы в единицу времени, равно его количеству, переходящему за это время из альвеол в кровь легочных капилляров. По аналогии – количество углекислого газа, поступающего в альвеолы из венозной крови, равно его количеству, удаляющемуся из альвеол в атмосферу.
Кислород переходит из альвеол в кровь легочных капилляров, а углекислота - в обратном направлении вследствие простого физического процесса диффузии; каждый из этих газов переходит из области более высокой его концентрации в область более низкой концентрации. Чрезвычайно тонкий альвеолярный эпителий не оказывает существенного сопротивления диффузии газов, и, поскольку в альвеолах концентрация кислорода обычно бывает выше, чем в крови, притекающий к легким по легочной артерии кислород диффундирует из альвеол в капилляры. Напротив, концентрация углекислоты в крови легочной артерии в нормальных условиях выше, чем в легочных альвеолах, и поэтому углекислота диффундирует из легочных капилляров в альвеолы.
В тканях всего тела, где происходит внутреннее дыхание, кислород переходит из капилляров в клетки, а углекислота - из клеток в капилляры путем диффузии. Вследствие непрерывного расщепления глюкозы и других веществ в клетках все время образуется углекислота и используется кислород. Поэтому концентрация кислорода в клетках всегда ниже, а концентрация углекислоты - выше, чем в капиллярах.
На всем своем пути от легких через кровь к тканям кислород движется из области с более высокой его концентрацией в область более низкой концентрации и, наконец, используется в клетках; углекислота движется из клеток, где она образуется, через кровь к легким и далее наружу - всегда по направлению к области с более низкой концентрацией.
33. Дыхательный и пневмотаксический центры, функциональные свойства дыхательных нейронов. Рефлекторная регуляция дыхания, рефлекс Геринга-Брейера. Вегетативные механизмы регуляции дыхания.Гуморальные механизмы регуляции дыхания, роль СО2.
Дыхательный центр – совокупность нервных клеток, расположенных в разных отделах цнс, обеспечивающих координированную ритмическую деятельность дыхательных мышц и приспособление дыхания к изменяющимся условиям внешней и внутренней среды организма.
Нервные клетки необходимые для ритмической деятельности дыхательных мышц в ретикулярной формации продолговатого мозга.
Бульбарный дыхательный центр. (в ретикулярной формации в области дна IV желудочка у нижнего угла ромбовидной ямки)
Инспираторная область включает в себя дорсомедиальную группу (ДМГ) нейронов и ростральную часть вентролатеральной группы (ВЛГ) нейронов. Она содержит:
Ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха (расположены в ВЛГ).
Полные инспираторные нейроны — эфферентные нейроны центра, иннервирующие мотонейроны мышц вдоха, активны в течение всего вдоха (расположены в ДМГ и ВЛГ).
Поздние инспираторные нейроны — эфферентные нейроны центра, иннервирующие мотонейроны мышц вдоха, активны в конце вдоха (расположены в ДМГ и ВЛГ).
Инспирация – растяжение легких и возбуждение эспираторных нейронов.
Экспираторная область включает в себя ВЛГ нейронов, преимущественно ее каудальную часть. Она содержит:
Постинспираторные нейроны — интернейроны, активные в первой половине выдоха, тормозят ранние инспираторные нейроны (расположены в ростральной ВЛГ).
Экспираторные нейроны — эфферентные нейроны центра, иннервирующие мотонейроны мышц выдоха, активны во второй половине выдоха, преимущественно при усиленном выдохе (расположены в каудальной ВЛГ).
Преинспираторные нейроны — интернейроны, блокирующие возбуждение экспираторных нейронов, способствуют смене выдоха на вдох.
Пневмотаксический центр моста.
Локализация: верхняя часть моста (медиальное парабрахиальное ядро).
Эффект: тормозит инспираторные нейроны, ограничивает длительность вдоха и повышает частоту дыхания.
Перерезка в нижней части моста приводит к удлинению вдоха, при дополнительной перерезке блуждающих нервов происходит апнейзис (глубокие длительные вдохи и короткие выдохи).
Рефлекторная регуляция дыхания.
Влияние с хеморецепторов на дыхательный центр.
Гуморальные факторы, участвующие в регуляции дыхания: Ро2, Рсо2 и рН крови:
стимулируют легочную вентиляцию гиперкапния, гипоксемия и ацидоз; уменьшают легочную вентиляцию гипокапния, гипероксия и алкалоз.
Периферические хеморецепторы:
локализуются в сосудах (особенно в артериях), тканях внутренних органов, при этом их концентрация максимальна в синокаротидной и аортальной зонах;
- имеют высокую чувствительность к изменению Po2 артериальной крови (особенно к его снижению), в меньшей степени реагируют на изменения Рсо2 и рН;
- имеют короткое латентное время действия на дыхательный центр (~ 5 с).
Центральные хеморецепторы (хеморецепторная зона дыхательного центра):
локализуются на переднебоковой поверхности продолговатого мозга и моста в виде трех пар скопления нейронов;
отличаются высокой чувствительностью к изменению рН (пороговые колебания рН 0,01) и Рсо2 в ликворе;
имеют длительное латентное время действия на дыхательный центр (~ 25 с).
Влияние с механорецепторов на дыхательный центр
С механорецепторов легких регулируется частота и глубина дыхания.
- Рецепторы растяжения легких (медленно адаптируются) расположены в гладких мышцах трахеи и бронхах, реагируют на увеличение объема легких при вдохе; с них возникает инспираторно-тормозящий рефлекс Геринга — Брейера (если дыхательный объем >1 л)(этот рефлекс запускается значительным объемом вдыхаемого воздуха. Развивающееся при этом увеличение объема легких усиливает импульсацию от рецепторов растяжения, находящихся в дыхательных путях, и приводит к прекращению вдоха)
-Ирритантные рецепторы расположены в эпителии бронхов, реагируют на быстрое изменение объема легких, на механические воздействия (пыль) и на пары химических веществ:
- это быстро адаптирующиеся рецепторы, порог раздражения у них выше, чем у рецепторов растяжения легких;
- с них формируются рефлексы кашля, сужения бронхов при выдохе, инспираторно-облегчающий рефлекс Геринга — Брейера при спадении легких, который укорачивает выдох и способствует смене его на вдох.
- Юкстаальвеолярные рецепторы (J-рецепторы) локализуются в интерстиции альвеол у капилляров, реагируют на давление жидкости в межклеточном пространстве легких, с них формируется одышка и торможение у мотонейронов (ограничение физической нагрузки).
- Проприорецепторы дыхательных мышц участвуют в компенсации дыхательных нагрузок:
локализуются в дыхательной мускулатуре, преимущественно в межреберных мышцах;
усиливают сокращение дыхательной мускулатуры при увеличении сопротивления дыханию, ослабляют — при уменьшении сопротивления дыханию.
Роль CO2
Углекислота возбуждает дыхательный центр не только непосредственно, но и рефлекторно – через хеморецепторы каротидной рефлексогенной зоны. Повыхение напряжения углекислоты в крови, перфузирующей изолированный каротидный синус, связанный с организмом только нервными волокнами, приводит к усилению дыхательных движений. При понижении напряжения углекислоты в крови, перфузирующий каротидный синус , дыхательные движения тормозятся.
Пищеварение и его значение. Типы пищеварения. Системные механизмы голода и насыщения. Аппетит.
Пищеварение – это процесс механического и ферментативного расщепления сложных органический веществ до простых. (гидролиз)
Значение:
Пищеварение является начальным этапом обмена веществ, которые поступают с пищей. Жиры, белки, углеводы, витамины, вода, минеральные соли и другие крупномолекулярные соединения распадаются на более мелкие и простые. Они всасываются слизистой оболочкой и поступают в кровь и лимфу. Отсюда они извлекаются клетками других органов и тканей организма, для потребления энергии на обновление, рост и работу.
Типы пищеварения:
Внутриклеточное (лизосомальное)
Полостное(внеклеточное)
Пристеночное
Также можно разделить на:
Аутолитическое(ферменты пищи)
Симбиотное(ферменты микрофлоры)
Собственное(ферменты организма)
Голод
Чувство голода проявляется в возникновении неприятных ощущений, локализуемых в области желудка, часто сопровождается тошнотой, нередко отмечается упадок сил и слабость, обычно возникает при пустом желудке, искл:булимия(патологическое чувство голода, наблюдаемое при нарушениях состояния некоторых отделов головного мозга, когда проявляется патологическая прожорливость) Ощущение голода связано с широкой областью нс – пищевой центр. В его составе клетки, расположенные в разных участках коры больших полушарий, в подкорковых ядрах и ретикулярной формации промежуточного мозга, в частности в гипоталамусе. Функция пищевого центра – регуляция пищевого поведения.
Центр голода локализован в латеральных ядрах гипоталамуса. Разрушение этих ядер вызывает афагию; стимуляция — гиперфагию.
2 теории физиологических механизмов голода:
1.Голод обусловлен снижением питательных веществ в крови — глюкозы, жирных кислот, аминокислот (теория голодной крови Павлова)
2.голод – местное чувство, зависящее от поступления в головной мозг импульсов от интерорецепторов пищеварительного тракта.
гормон гастрин вырабатывает чувство голода:повышается секреция HCl, тормозит переход пищевого комка(химус), вырабатывается самостатин:тормозит секрецию гастрина, вырабатывается гистамин – увеличивает проницаемость кр.сосудов, капилляров, расширяет кровеносный сосуды. (теория пустого желудка Кеннон).
Аппетит — активатор пищевого поведения.
Аппетит индивидуально вырабатывается и отражает не столько потребность в пище вообще, сколько потребность в связи со спецификой обмена веществ и дефицитом тех или иных компонентов пищи, индивидуальных и групповых привычек, особенностью продуктов, которым отдается предпочтение.
- Избирательный аппетит - это влечение человека к определенному роду пищи, чаще тому, который содержит недостающие организму вещества
Расстройства аппетита: анорексия, булемия, извращение аппетита.
Насыщение.
Центр насыщения локализован в вентромедиальных ядрах гипоталамуса.Разрушение этих ядер вызывает гиперфагию, стимуляция — афагию.
Стимулируют центр насыщения некоторые вещества крови (глюкоза, аминокислоты, жирные кислоты, кислоты цикла Кребса), повышение температуры крови, пептидный гормон жировой ткани — лептин.
Виды (фазы) насыщения.
- Сенсорное насыщение (предрезорбционное, первичное).Формируется во время приема пищи с рецепторов полости рта, пищевода, желудка.
- Центр насыщения стимулируется как рефлекторным путем, так и гуморально в результате мобилизации депонированных питательных веществ (гликогена, липидов).
- Позволяет окончить прием пищи еще до всасывания питательных веществ из тонкого кишечника.
- Обменное насыщение (пострезорбционное, вторичное)
- Формируется в результате всасывания питательных веществ в кровь.
-Стабилизируется уровень питательных веществ в крови и пролонгируются промежутки между приемами пищи.
-Восстанавливаются депо питательных веществ в организме.
Голод, насыщение, аппетит являются средствами оценки пищевой мотивации и приема необходимого количества питательных веществ.
Пищеварение в ротовой полости, прием пищи, вкусовая рецепция, глотание. Слюнные железы, состав и свойства слюны. Регуляция слюноотделения.
Функции пищеварения:
Захват и механическая переработка пищи в процессе жевания.
Происходит частичная ферментативная обработка
Формирование пищевого комка и его продвижение
Вкусовая и температурная рецепция (участвует в формировании чувства голода и насыщения)
Всасывание некоторых растворенных в воде веществ.
Анализ вкусовых свойств пищи и разделение их на пищевые и отвергаемые. (Участвуют зрительный, обонятельный, тактильный, температурный анализаторы и кортико-лимбические структуры).
Механическая обработка пищи — жевание(прием пищи)
Роль жевательных мышц:
-Главные мышцы осуществляют основные жевательные движения: опускание нижней челюсти (двубрюшная, подбородочно-подъязычная, челюстно-подъязычная), ее поднимание и смыкание зубов (жевательная, височная, крыловидная мышцы).
-Вспомогательные мышцы (языка, щек, губ) удерживают пищу между жевательными поверхностями.
Зубы осуществляют откусывание, разламывание, раздавливание и растирание пищи.
Рефлекторный акт жевания является сложным ритмическим безусловным рефлексом с хорошо выраженным произвольным контролем.
-Афферентное звено рефлекса осуществляется с тактильных, вкусовых, температурных рецепторов слизистой оболочки рта и проприорецепторов аппарата жевания. Оно обеспечивается в основном сенсорными волокнами и ядрами V нерва.
-Центр жевания (центральный генератор ритма жевания) находится в ретикулярной формации продолговатого мозга и моста. Он вызывает ритмическое возбуждение мотонейронов мышц, поднимающих и опускающих нижнюю челюсть.
-Основное эфферентное влияние центра жевания осуществляется через двигательное ядро V нерва.
-Мастикациография — метод исследования биомеханики жевательного акта, позволяет зарегистрировать 5 фаз: 1) покой; 2) введение пищи в рот; 3) ориентировочная; 4) основная; 5) формирование пищевого комка.
Глотание — переход пищевого комка из полости рта в желудок.
- В процессе передвижения пищевого комка из полости рта в пищевод происходит последовательное возбуждение рецепторов корня языка, мягкого нёба, глотки и пищевода. Импульсация по чувствительным волокнам IX и X черепных нервов поступает в центр глотания.
- Центр глотания, расположенный в продолговатом мозге и мосту, включает в себя чувствительное ядро одиночного пути и двойное (двигательное) ядро IX, X нервов и прилегающие зоны ретикулярной формации.
- В результате этого обеспечивается строго координированная последовательность сокращения мышц, участвующих в акте глотания: челюстно-подъязычных, языка, мягкого нёба, глотки, гортани, надгортанника и пищевода.
- Центр глотания функционально связан с центрами жевания и дыхания: рефлекс глотания останавливает акт жевания и дыхание (обычно в фазе вдоха).
Фазы акта глотания
Ротовая (произвольная) приводит к формированию пищевого комка, который выносится на корень языка.
Глоточная сопровождается напряжением мягкого нёба, подъемом гортани и опусканием надгортанника, в результате комок пищи перемещается к началу пищевода.
Пищеводная.
- Передвижение пищи в пищеводе осуществляется с помощью перистальтики
- Парасимпатические влияния стимулируют перистальтику пищевода, сокращают верхний (поперечнополосатый) пищеводный сфинктер и расслабляют нижний (гладкомышечный) пищеводный сфинктер (НПС).
- Симпатические влияния тормозят перистальтику пищевода (3-адренорецепторы) и повышают тонус НПС (а-адренорецепторы).
Слюнные железы:3 пары крупных и множество мелких (подъязычная, поднижнечелюстная, околоушная)
Количество и состав слюны.
В сутки секретируется 1,5—2,0 л слюны. рН 5,8—7,4.
Слюна содержит электролиты (К+, Са2+, НС03), мочевину, имеется а-амилаза и низкая активность других ферментов (липазы, протеазы, нуклеазы и др.); гликопротеин муцин; хорошая бактерицидная активность за счет лизоцима, иммуноглобулинов (особенно Ig А), лейкоцитов.
Физиологическое значение слюны: участвует в увлажнении и ослизнении пищевой массы, формировании пищевого комка, переваривании углеводов, возникновении вкусовых ощущений, необходима для устной речи.
Регуляция слюноотделения осуществляется преимущественно нервными механизмами по типу безусловных и условных рефлексов.
- Важнейшим рецепторным полем являются вкусовые, тактильные, болевые, температурные рецепторы полости рта.
- Афферентная импульсация по волокнам V, VII, IX и X черепных нервов поступает в продолговатый мозг, где переключается на бульбарно-спинальный центр слюноотделения. Он включает в себя верхнее и нижнее слюноотделительные ядра продолговатого мозга, иннервирующие слюнные железы парасимпатическими волокнами VII и IX нервов, и нейроны сегментов Th, — Th3, осуществляющих симпатическую иннервацию.
- Парасимпатические влияния (ацетилхолин, М-холинорецепторы) приводят к выделению большого количества жидкой слюны с высокой концентрацией электролитов и низкой — муцина.
- Симпатические влияния через 3-адренорецепторы стимулируют секрецию ферментов слюны (слабый эффект через а-адренорецепторы — секрецию воды, Na+, Сl-).
Пищеварение в желудке. Моторная и секреторная деятельность желудка, их регуляция. Железы желудка, состав и свойства желудочного сока.
В желудке происходит депонирование пищи. Имеет 3хслойною мышечную оболочку, происходит механическая обработка пищи, ее дальнейшее распределение. Играет важную роль в регуляции голода и насыщения, происходит частичное ферментативное переваривание. Также за счет соляной кислоты защита от бактерий, может происходить частичное всасывание в-в, Играет роль в регуляции пищеварения в кишечнике. растворенных в воде. Выделает желудочные гормоны.
Моторная функция:
Сокращение гладких мышечных волокон стенки желудка обеспечивают моторную функцию. Значение ее:перемешивание содержимого желудка и передвижение пищи из желудка в кишку. Функция резервуара – проксимальный желудок(дно и часть тебя желудка)
Натощак объем желудка 50мл, максимальный объем 1500мл.
-Релаксация «проксимального» желудка во время пищи – симпатические влияния (через бета-адренорецепторы), вагусные нехолинергические влияния (медиатор ВИП), гастрин, ГИП.
-Сокращения – вагусные холинергические влияния.
Перемешивание передвижение пищи – дистальный желудок(тело и андрально-пилорический отдел)
Виды движений желудка:
-перистальтика
-тонические волны желудка
-систолические сокращения антрально-пилорического отдела приводят к эвакуации химуса в 12-перстную кишку, а также измельчение химуса, отбрасывание назад нерастертых частиц.
В регулировании пищи в кишечник важная роль принадлежит пилорическому сфинктеру, расположенному в конце пилорического отдела желудка и закрывающему выходное отверстие желудка, и препилорическому сфинктеру, имеющемуся между фундальной и пилорической частью желудка.
Типы сокращений мускулатуры желудка:
Первый наблюдается после приема пищи, когда происходит секреция кислого желудочного сока. Частота сокращений 5-6 в минуту.После 1-2 часа сокращения могут отсутствовать.
При переходе пищи из желудка в кишку – во время эвакуации пищи из желудка, а также при пустом желудке во время периодической моторной его деятельности. Он характеризуется более медленными колебаниями давления большой амплитуды. Сокращения 2го типа слабее в области фундальной части желудка и в 2-3 раза сильнее в пилорической части.
Продолжительность волны сокращения мускулатуры желудка от 10 до 30 секунд. В обычных условиях пищеварения сокращения желудка возникают в результате механического раздражения его стенки пищей.
Регуляция моторики желудка:
-стимулируют моторику желудка: парасимпатические влияния (ацетилхолин), гастрин, мотилин и т.д.
- тормозят: симпатические влияния и адреналин, ГИП, соматостатин, секретин, серотонин и др.
Секреторная деятельность:
Регуляция желудочной секреции, фазы:
Энцефалическая (мозговая) фаза (запах пищи, запальный сок)
Желудочная (в желудок попадает пища)
Кишечная
СНС снижает секрецию, ПСНС повышает.
В желудке вырабатывается желудочный сок. За сутки около 2х литров. pH=1,5-2ед. рН регулируется соляной кислотой. Он вырабатывается железами, которые состоят из железистых клеток. Выделяют 4 типа: главные- пепсиногены – пепсин – белки – пептиды (неактивные ферменты, гидролизирующие белки, расщепляющие молекулу на пептиды)Гастриксин преобразует белки молока (сворачинание) – казеиноген, затем казеин.
Обкладочные клетки синтезируют HCl.
Механизм секреции HCl: источником H+ является диссоциация Н2СО3; Н+ переносится в просвет желудка с помощью Н+-насоса, Сl- в результате ионообмна на НСО3-.
Свойства соляной кислоты:
-Активирует пепсиногены, отщепляя С-концевой ингибирующий пептид
-Устанавливает оптимум рН для действия пепсина
-вызывает денатурацию и набухание белков
-Оказывает бактерицидный эффект
Поджелудочная железа, состав и свойства поджелудочного сока, регуляция секреции. Печень и ее функции. Состав и свойства желчи, ее роль в процессах пищеварения.
В паренхиме железы выделяют экзокринную и эндокринную части. Экзокринная часть вырабатывает основные пищеварительные ферменты, имеет главный и добавочный выводные протоки, которые открываются на большом и малом дуоденальных сосочках. Эндокринная часть железы образует островковый аппарат (островки Лангерганса), вырабатывает инсулин и глюкагон.
Основные ферменты панкреатического сока.(2-2,5л за сутки, рН около 8 ед.)
Протеазы (переваривают белки):
-трипсин, трипсин-ингибитор защищает поджелудочную железу от самопереваривания;
-химотрипсин;
-карбоксипептидазы А и В
- Липазы (переваривают липиды)
- Нуклеазы (переваривают нуклеиновые кислоты).ДНКазы, РНКазы – расщепляют ДНК и РНК на нуклеотиды.
Также содержит ферменты, расщеплющие жиры – липаза, фосфолипаза А2, расщепляет углеводы – альфа-амилаза. Играют важную роль в полостном пищеварении.
Регуляция панкреатической секреции.
Фазы отделения поджелудочного сока:
1) мозговая (менее выражена, чем в желудке);
2) желудочная;
3) кишечная (основная).
Нервные влияния (менее выражены, чем гуморальные):
-парасимпатические стимулируют секрецию ферментов и НС03- (М-холинорецепторы ИФ3\ДАГ Са2+↑)
- симпатические тормозят секрецию через бетта-адренорецепторы (са2+↓)
- пептидергические (стимулирующие и тормозные).
Гуморальные влияния на панкреатическую секрецию:
- стимулируют: секретин (секреция Н20, НСО3-), ХЦК-ПЗ (секреция ферментов), гастрин (а также серотонин, инсулин, бомбезин);
- тормозят: соматостатин, ГИП, глюкагон;
Приспособительная секреция поджелудочного сока на различные виды пищи и пищевые рационы.
Печень - крупная пищеварительная железа, вырабатывающая желчь; функции: участвует во всех видах обмена веществ, осуществляет детоксикацию и пр. Структурно-функциональной единицей печени является сосудистая долька. Печеночные клетки группируются в дольке в ряды (балки), расположенные радиально.
Желчесекреторная функция печени.
Количество и состав печеночной желчи (~ 1 л/сут, рН 8,2; НС03-).
- Желчные кислоты: в печени из холестерина образуются холевая и хенодезоксихолевая кислоты.
- Холестерин
- Билирубин
- Фосфолипиды (особенно лецитин).
Функции желчи.
- Эмульгирует жиры с образованием мицелл.
- Повышает панкреатическую секрецию и активность липазы.
- Способствует всасыванию липидов и жирорастворимых витаминов.
- Стимулирует образование и выделение желчи.
- Инактивирует пепсины.
- Обладает бактерицидным действием.
Образование желчи (холерез)
- Секреция (активный транспорт) из гепатоцита в желчный капилляр желчных кислот, холестерина, билирубина, электролитов (Na+,Cl- , НС03-), по осмотическому градиенту — воды.
- Печеночно-кишечная циркуляция желчных кислот.
В сутки по кругу "печень -> желчь -> тонкий кишечник —>реабсорбция в конце подвздошной кишки -> портальная кровь -> печень" циркулирует ~ 24 г желчных кислот.
Вновь образуется в печени 0,5 г желчных кислот в сутки и столько же покидает организм через кишечник.
Желчевыделение (холекинез).
Желчевыводящие капилляры, ходы и протоки (общий печеночный, пузырный, общий желчный), их сфинктеры.
Функции желчного пузыря.
- Резервуарная функция (60—80 мл желчи, в патологических условиях до 1000 мл).
- Концентрирующая: за счет реабсорбции Na+, Сl- и воды концентрация желчных кислот и холестерина увеличивается в 8—10 раз.
-Выделение желчи (сокращение протоков, пузыря, расслабление сфинктеров).
Стимулируют: парасимпатические влияния, ХЦК-ПЗ, гастрин, липиды пищи, сульфат магния;
Тормозят: симпатические влияния, панкреатический пептид, глюкагон.
Пищеварение в тонком кишечнике, моторная и секреторная функции тонкого кишечника, их регуляция. Состав и свойства кишечного сока. Полостное и пристеночное пищеварение. Мембранное всасывание воды, минеральных солей и органических соединений в тонком кишечнике.
Переваривание:
12-перстная кишка
Тощая
Подвздошная
Происходят процессы ферментативной обработки, процессы всасывания, регуляция пищеварительных процессов.
Секреция.
Осуществляется нервным и гуморальным путем. Симпатика снижает секрецию и моторику, парасимпатика увеличивает. Выделяется холецистокинин (панкреозилин) Стимулирует образование желчи, панкреалического сока, карбонатов и трипсиногенов. Трипсин оказывает отрицательное влияние на секрецию холицистокинина.
Моторика.
Перемешивание химуса с помощью мелкой перистальтики, ритмической сегментации, маятникообразного движения и сокращения ворсинок.
Передвижение химуса с помощью глубокой перистальтики. Химус в кишке 4-6 часов.
Эвакуация химуса в толстый кишечник при рефлекторном открывании илеоцекального сфинктера.
Регуляция моторики:
- стимулируют парасимпатика, гастрин, мотилин, грубая и жирная пища
- тормозят: симпатические влияния, адреналин.
Состав и свойства кишечного сока.
Количество – 2л\сут, рН=8
Ферментный состав:
- протеазы, дипептидазы, аминопептидаза
- энтерокиназа (активирует трипсиноген)
- карбогидразы: мальтаза, лактаза, сахараза
- фосфатаза, нуклеаза и пр.
Секреция желчной слизи, содержащей гликопротеины.
Регуляция секреции:
Особенность: не выражена стимуляция во время приема пищи
Стимуляция секреции местным механическим и химическим (HCl, продукты белкового и липидного гидролиза) раздражителями химуса.
Рефлексы метасимпатической нервной системы (медиаторы: ВИП, ацетилколин и др.) и центральные вегетативные рефлексы.
Полостное пищеварение:
осуществляется за счет пищеварительных секретов и их ферментов, поступивших в полость тонкой кишки (секрет поджелудочной железы, желчь, кишечный сок). В результате гидролизуются крупномолекулярные вещества и образуются в основном олигомеры. Последующий их гидролиз происходит в зоне, прилегающей к слизистой оболочке, и непосредственно на ней. Из полости тонкой кишки вещества поступают в слой кишечной слизи.
Пристеночное пищеварение:
Это соприкосновение пищи со слизистой оболочкой, очень большая пористая поверхность тонкой кишки способствует усилению энзиматических процессов, адсорбируя ферменты и являясь своеобразным пористым катализатором. Окончательное расщепление пит.в-в происходит на той же поверхности тонкой кишки, которая обладает функцией всасывания. осуществляется в трех зонах: в слое слизи, гликокаликсе и на апикальных мембранах энтероцитов. Образовавшиеся в результате пищеварения мономеры всасываются в кровь и лимфу.
Всасывание воды:
- вода может диффундировать в обе стороны, поэтому кишечный химус изотоничен плазме крови.
- при всасывании коносахаридов, аминокислот, Na+, вода следует за ними в соответствии с осмотическим градиентом.
Всасывание продуктов гидролиза белков (аминокислоты и часть дипептидов).
- В апикальной мембране (щеточной каемке) имеется 5 видов переносчиков аминокислот (для кислых, основных, нейтральных кислот, иминокислот, Р- и у-аминокислот).
- На апикальной мембране происходит активный транспорт с использованием градиента Na+, на базальной мембране — облегченная диффузия.
- Часть дипептидов всасывается (с использованием переносчиков и энергии градиента Н+) и гидролизуется в ли-зосомах.
Всасывание продуктов гидролиза углеводов.
- Всасываются только моносахариды с использованием активного транспорта (энергия градиента Na+), облегченной и простой диффузии.
Глюкоза, галактоза через апикальную мембрану переносятся путем активного транспорта, через базальную мембрану переходят путем простой и облегченной диффузии.
Фруктоза транспортируется путем облегченной диффузии, манноза и пентоза — простой диффузии.
Регуляция всасывания углеводов:
Усиливают парасимпатические влияния, глюкокортикоиды, тироксин, инсулин.
Тормозят симпатические влияния, соматостатин.
Всасывание продуктов гидролиза жиров.
Мицеллы свободно проникают в просветы между микроворсинками и достигают поверхности энтероцита.
Липидное содержимое мицеллы (жирные кислоты, глицерол, 2-моноацилглицерол, продукты переваривания фосфолипидов и эфиров холестерина) диффундируют с помощью переносчиков (возможно, активным транспортом) внутрь энтероцита .
В гранулярной ЭПС энтероцитов происходит ресинтез триглицеридов, фосфолипидов и холестеролэфиров, которые в аппарате Гольджи упаковываются в липопротеиновую оболочку, образуя хиломикроны и липопротеины очень низкой плотности (d ~ 50 нм), которые транспортируются из энтероцита путем экзоцитоза.
Затем хиломикроны и ЛПОНП поступают в лимфатические сосуды кишечника.
Короткоцепочечные жирные кислоты (< 10 атомов углерода) и глицерол могут всасываться вне состава мицелл и поступать в кровь.
Регуляция всасывания липидов.
Усиливают парасимпатические влияния, секретин, ХЦК-ПЗ, глюкокортикоиды.
Тормозят симпатические влияния, адреналин.
В норме всасывается около 98% липидов пищи.
Всасывание ионов.
Поступление Na+ в энтероцит через апикальную мембрану происходит по электрохимическому градиенту с участием переносчика, из энтероцита через базальную и латеральную мембраны с помощью Na+, К+-насоса (всасывается в сутки ~ 25 г Na+).
Всасывание Сl- совпадает (через потенциал) с транспортом Na+, но Сl- всасывается больше, так как есть еще обмен Сl- на НС03-
Всасывание Са2+ происходит с помощью Са2+-связывающего белка и витамина D3, паратгормон активирует всасывание. Выход Са2+ из энтероцита происходит с участием Са2+\Na+ - ионообменника и Са2+ - насоса.
Пищеварение в толстом кишечнике. Секреторная функции толстого кишечника, симбионтное пищеварение в толстом кишечнике. Моторная функция толстого кишечника, дефекация.
В нем происходит формирование каловых масс, симбиотическое расщепление клетчатки с участием микрофлоры, всасывание воды и растворенных веществ (0,5л воды), с калом выходит около 100 мл воды, остальное всасывается, происходит синтез витаминов (К, некоторый витаминов группы В). Бактерии: бифидобактерии, бактероиды – главная микрофлора, лактобактерии, стрептококки, грибы. Микрофлора образуется в подвздошной части кишечника и в толстом кишечнике.
Секреторная функция
Количество и состав кишечного сока (0,05 л/сут, рН 8,7).
Малая активность ферментов (пептидазы, липазы, амилазы, нуклеазы, щелочной фосфатазы).
Большое количество слизи (бокаловидные клетки).
Регуляция секреции.
Основной стимул — механическое раздражение.
Преобладают местные механизмы регуляции: рефлексы метасимпатической нервной системы, секреция ВИП, энтерогастрона, серотонина.
Моторная функция толстого кишечника.
Питательные вещества принятой пищи начинают поступать в толстый кишечник через 6—7 ч, заполняют его через 24 ч (~ 1500 мл химуса в сутки), полное опорожнение происходит за 2—3 дня.
- Резервуарная функция обеспечивается гаустрацией, тонусом сфинктеров, пластическим тонусом.
- Функция перемешивания содержимого обеспечивается гаустрацией, маятникообразными движениями, мелкой перистальтикой и антиперистальтикой.
- Функция передвижения химуса осуществляется в результате глубокой пропульсивной перистальтики, возникающей 3—4 раза в сутки. Снижение тонуса сигмы способствует переходу кала в прямую кишку, повышение тонуса ведет к запорам.
Регуляция моторики толстого кишечника.
Стимулируют моторику механические и химические раздражители химуса (напр., пищевые волокна) через парасимпатические влияния. Тормозят моторику симпатические влияния, серотонин, адреналин.
Акт дефекации (норма — от 2 раз в сутки до 1 раза в двое суток).
Раздражение механорецепторов прямой кишки (давление 30—40 мм рт. ст.), афферентная импульсация по волокнам тазового и срамного нервов.
Центральное звено — аноспинальный центр (S2_4), а также моторные центры диафрагмы и брюшных мышц.
Эфферентное звено — парасимпатические волокна тазового нерва, соматические волокна диафрагмальных, межреберных, подвздошно-подчревных и подвздошно-паховых нервов.
Эффекторное звено:
- расслабление внутреннего анального сфинктера (гладкая мышца, непроизвольный контроль);
- расслабление наружного анального сфинктера (поперечно-полосатая мышца, произвольный контроль с 2 лет);
- перистальтика толстой кишки;
- сокращение мышц брюшного пресса и диафрагмы.
Симбионтное пищеварение — гидролиз питательных веществ за счет ферментов, синтезированных симбионтами макроорганизма — бактериями и простейшими пищеварительного тракта. У человека клетчатка пищи по типу собственного пищеварения из-за отсутствия соответствующего фермента в секретах желез не гидролизуется поэтому переваривание ее ферментами симбионтов в толстой кишке является важным процессом. В результате симбионтного пищеварения образуются вторичные пищевые вещества в отличие от первичных, образующихся в результате собственного пищеварения. Аутолитическое пищеварение осуществляется за счет экзогенных гидролаз, которые вводятся в организм в составе принимаемой пищи. Роль данного пищеварения существенна при недостаточно развитом собственном пищеварении.
Характеристика обмена веществ и энергии, анаболизм и катаболизм. АТФ как универсальный источник энергии, этапы синтеза АТФ в организме. Энергетическая ценность белков, жиров и углеводов. Калориметрический эквивалент кислорода. Дыхательный коэффициент.
Обмен веществ — характерный признак жизни. Он обеспечивает непрерывное образование, обновление, разрушение клеточных структур, синтез и разрушение различных химических соединений. Включает в себя два процесса.
Анаболизм (ассимиляция) — усвоение организмом питательных веществ, в результате которого они становятся частью биологических структур или откладываются в виде депо (сопровождается ростом и увеличением массы тела).
Катаболизм (диссимиляция) — образование в организме из сложных веществ более простых веществ, с использованием их преимущественно для энергетических целей (сопровождается снижением массы тела).
Молекула АТФ – основное универсальное вещество, обладающее энергией(в большом количестве не может бвть запасено)
Происходит ресинтез АТФ из АДФ+Ф и из АТФ АДФ+Ф
АТФ образуется из:
Энергетическая ценность белков, жиров и углеводов.
Количество энергии, выделяемой при сгорании какого-либо вещества, не зависит от этапов его распада. Известно, что углеводы и белки дают в среднем около 17,16 кДж/г (4,1 ккал/г) энергии. Самой высокой энергетической способностью обладают жиры: 1 г жира дает 38 гДж/г (9,1 ккал/г) энергии, что больше количества энергии, выделяемой при окислении белков и углеводов, вместе взятых.
Отношение объема выделенного углекислого газа к объему поглощенного организмом кислорода называется дыхательным коэффициентом. По величине последнего можно судить о характере окислительных веществ в организме.
Методы исследования обмена веществ: прямая и непрямая калориметрия. Основной и валовый обмены. Условия измерения и нормальные величины основного обмена человека. Зависимость интенсивности основного обмена от массы и площади поверхности организмов, правило поверхности.
Для определения интенсивности обмена веществ и энергии используют прямые и непрямые методы калориметрии. Метод прямой калориметрии основан на непосредственном определении тепла, выделяемого в процессе жизнедеятельности организма. Для этого человека помещают в специальную калориметрическую камеру, в которой учитывается все количество тепла, отдаваемого телом человека. Метод сложен и применяется только в научно-исследовательских учреждениях. На практике чаще используют метод непрямой калориметрии. Суть его заключается в том, что вначале определяют объем легочной вентиляции, а затем количество поглощенного кислорода и выделенного углекислого газа.(камерный(закрытый) метод и некамерные(открытый, полуоткрытый, закрытый методы)
Валовый обмен
Об интенсивности обмена веществ в целом {валового, или общего, обмена) и характере веществ, окисляющихся в организме, можно судить по объему потребленного кислорода и продуктам распада, выделяемым из организма. Так, количество распавшегося в организме белка определяют по количеству азота, выделенного с мочой. Количество окисленных углеводов и жиров - по объему выделенной углекислоты и потребленного за то же время кислорода, т. е. по данным газообмена. При этом учитывают, что углекислый газ образуется в организме не только при окислении жиров и углеводов, но и при окислении белков. Определив по количеству выведенного из организма азота количество белка, распавшегося в организме за сутки, можно высчитать содержание в нем углерода (в белке содержится в среднем 52% углерода). Разность между всем количеством углерода, входившего в состав распавшегося в теле белка, и количеством углерода, содержащегося в моче (в последней углерод содержится преимущественно в продуктах распада белка), дает возможность рассчитать, сколько углерода, находившегося в белке, пошло на образование углекислого газа и сколько для этого потребовалось кислорода. Вычтя из общего количества кислорода, поглощенного организмом за время исследования, количество кислорода, затраченного на окисление белков, узнают, сколько кислорода использовано на окисление жиров и углеводов. Дальнейший расчет позволяет узнать количество кислорода, затраченного на образование отдельно углеводов и отдельно жиров, а отсюда определить количество тех и других, окислившихся в организме. Основанием для расчета служит то обстоятельство, что при окислении 1 г углеводов и 1 г жиров используются разные количества кислорода и выделяются разные количества углекислого газа.
Основной обмен (ОО) — это минимальный уровень обмена, который определяется при стандартных условиях: утром, в покое (лежа), натощак, при температуре комфорта (18—20 °С).
Физиологическое значение — поддержание минимально необходимого для жизни клеток уровня окислительных процессов и деятельности постоянно работающих органов и систем (сердце, почки, дыхательные мышцы, тонус сосудов и др.).
Величина
ОО определяется
по формуле
Дрейера (ОО =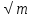 \K
* возраст0,1333,
где К = 0,1015 у мужчин; 0,1029 у женщин).
\K
* возраст0,1333,
где К = 0,1015 у мужчин; 0,1029 у женщин).
Нормальные величины ОО
Факторы, определяющие величину ОО:
- Пол (у женщин на 10 % ниже, чем у мужчин).
- Возраст (у детей относительно выше, чем у взрослых, в пожилом возрасте снижается).
- Уровень гормонов в крови (при избытке тироксина и адреналина увеличен, при недостатке снижен).
Для мужчины среднего возраста (примерно 35 лет), среднего роста (примерно 165 см) и со средней массой тела (примерно 70 кг) основной обмен равен 4,19 кДж (1 ккал) на 1 кг массы тела в час, или 7117 кДж (1700 ккал) в сутки. У женщин той же массы он примерно на 10 % ниже
Правило поверхности
Если пересчитать интенсивность основного обмена на 1 кг массы тела, то окажется, что у теплокровных животных разных видов и у людей с разной массой тела и ростом она весьма различна. Если же произвести перерасчет интенсивности основного обмена на 1 м2 поверхности тела, полученные у разных животных и людей величины различаются не столь резко
Правило поверхности — затраты энергии теплокровными животными пропорциональны величине поверхности тела (у мужчины примерно 4000 кДж/м2). Чем меньше площадь поверхности, тем больше теплообмен.
S=L2, M=L3 при увеличении массы площадь непрямолинейно возрастает (на 2\3).
Особенности обмена энергии при умственном и физическом труде. Возрастные и профессиональные особенности энергообмена. Восполнение энергозатрат питанием. Нормы питания, составление пищевого рациона.
Обмен энергии при физическом труде
Мышечная работа увеличивает расход энергии, суточный расход энергии при физическом труде превышает величину основного обмена.
При мышечной работе освобождается тепловая и механическая энергия. Отношение механической энергии ко всей энергии, затраченной на работу, выраженное в процентах, называется коэффициентом полезного действия. При физическом труде человека КПД в среднем 20 %. КПД изменяется в зависимости от условий( у нетренированных людей он ниже, чем у тренированных, и увеличивается по мере тренировки).
Затраты энергии тем больше, чем интенсивнее мышечная работа. Степень энергетических затрат определяется коэффициентом физической активности (КФАУчитывая особенности прифессии все мужское население разделено на 5 групп.
I группа - работники преимущественно умственного труда, очень легкая физическая активность
II группа - работники, занятые легким трудом, легкая физическая активность
III группа - работники средней тяжести труда, средняя физическая активность
IV группа - работники тяжелого физического труда, высокая физическая активность
V группа - работники особо тяжелого физического труда, очень высокая физическая активность
Значительные различия энергетической потребности в группах зависят от пола (у мужчин больше), возраста (снижаются после 40 лет), степени активности отдыха и уровня коммунального обслуживания.В старости энерготраты снижаются и к 80 годам составляют 2000—2200 ккал.
Обмен энергии при умственном труде
При умственном труде энерготраты значительно ниже, чем при физическом.
формы умственного труда, если они не сопровождаются движением, вызывают ничтожное (2—3 %) повышение затраты энергии по сравнению с полным покоем. Однако в большинстве случаев различные виды умственного труда сопровождаются мышечной деятельностью, в особенности при эмоциональном возбуждении работающего, поэтому и энерготраты могут быть относительно большими. Пережитое эмоциональное возбуждение может вызвать в течение нескольких последующих дней повышение обмена на 11—19 %.
Питание — процесс поступления, переваривания, всасывания и усвоения в организме пищевых веществ, необходимых для покрытия энергетических и пластических потребностей организма, образования его физиологически активных веществ.
Энергия может храниться в форме гликогена(в печени, мышцах), он они ограниченны, так как гликоген требует большое кол-во воды. 1 кг гликогена = 1кг воды, поэтому более эффективно хранение в форме жиров.
Нормы питания.
Суточная потребность организма в белках, углеводах, жирах, минеральных солях, витаминах.
Потребность организма в питательных веществах зависит от географических и климатических условий, от возраста, профессии, состояния организма (усиленный рост, беременность, период лактации).
Основные принципы рационального питания.
Энергетическая ценность питания должна соответствовать энергетическим затратам организма.
Состав питательных веществ должен соответствовать физиологическим потребностям в них организма.
Питание должно быть разнообразно, содержать свежие натуральные продукты растительного и животного происхождения.
Суточное количество пищи должно поступать в организм порционно (регулярность, кратность и чередование приемов пищи).
При работе средней тяжести требуется около 120г белка, при тяжелой – 150-160г. Не менее 30% этого белка должно быть животного происхождения. Жиров 100г, так как в их состав входят жирорастворимые витамины и липоиды, необходимые для построения клеток. Суточное количество углеводов – 400-500г.
Характеристика обмена белков, незаменимые аминокислоты. Азотистый баланс. Регуляция белкового обмена.
Обмен белков составляет 15-20% массы тела.
Функции обмена:
Ферментативная ферменты по хим.природе – белки.
Пластическая – белки мембран, коллаген, эластин, кератин и т.д.
Транспортная – перенос с помощью белков билирубина, липидов, кислорода, железа и др.
Энергетическая – на долю белка приходится 12-18% энергообеспечения(окисление 1г белков = 4ккал)
Гормональная – белково-пептидные гормоны составляют около 80% всех гормонов.
Сократительная – актин и миозин.
Защитная – иммуноглобулины, интерферон, фибриноген, плазмин.
Регуляция работы генов – факторы транскрипции.
В сутки человеку необходимо 100г белка. Белки характеризуется аминокислотным составом. Полноценные содержат все аминокислоты.Около половины аминокислот (8 из 20) не могут синтезироваться в организме (незаменимые аминокислоты), они поступают с пищей:
Лизин, лейцин, изолейцин, валин, фенилаланин, трионин, триптофан, метионин.
Азотистый баланс — соотношение количества азота, поступившего с пищей (белки) и выделенного из организма(мочевина, мочевая кислота, креатинин и др.) Так как 1 г азота содержится в 6,25 г белка, то, зная количество выделенного азота с мочой, можно определить количество усвоенного белка.
Азотистое равновесие (нейтральный баланс азота): у взрослого человека при адекватном питании количество введенного азота с пищей равно выведенному; при увеличении потребления белка устанавливается на новом, более высоком уровне.
Положительный азотистый баланс: синтез белка преобладает над распадом (при увеличении массы тела, во время беременности, в период роста организма, при усиленных тренировках в связи с ростом мышечной массы).
Отрицательный азотистый баланс: количество выведенного азота больше, чем поступающего с пищей (белковое голодание, питание неполноценными белками, различные заболевания).
Нейрогуморальная регуляция обмена белков.
Синтез белка стимулируют СТГ(гормон роста), тироксин и трийодтиронин, глюкокортикоиды в печени.
Распад белка увеличивают глюкокортикоиды (в мышцах и лимфоидной ткани) и глюкагон.
Характеристика обмена углеводов. Роль гликогена в энергообеспечении организма. Регуляция углеводного обмена.
Обмен углеводов (составляют 0,6 % массы тела, суточная потребность 300—350 г).
Функции углеводов:
1) энергетическая — основной источник энергии в организме, углеводы обеспечивают не менее 50 % суточного энергообеспечения (окисление 1 г углеводов освобождает 4 ккал);
2) образуют депо (в виде гликогена) легко мобилизируемого энергетического материала;
3) пластическая — продукты промежуточного обмена углеводов используются для синтеза аминокислот, липидов, полисахаридов;
4) углеводы, входящие в гликопротеиды, определяют их видовую и тканевую специфичность.
Роль гликогена в энергообеспечении организма.
Гликоген откладывается также в мышцах, где его содержится около 1—2%. Количество гликогена в мышцах увеличивается при обильном питании и уменьшается во время голодания. При работе мышц происходит усиленное расщепление гликогена. Под влиянием фермента фосфорилазы, которая активируется в начале мышечного сокращения, к гликогену присоединяются молекулы фосфорной кислоты, и он расщепляется до глюкозо-1-фосфата, являющегося одним из источников энергии мышечного сокращения.
Нейрогуморальная регуляция обмена углеводов.
Активирует использование глюкозы (снижает ее уровень в крови) инсулин, он способствует использованию глюкозы в тканях (особенно мышечной и жировой) и депонированию ее в виде гликогена (особенно в печени и мышцах) и синтезу аминокислот (белков). Парасимпатические влияния (М-холинорецепторы) и симпатические влияния ((3-адренорецепторы) стимулируют секрецию инсулина, симпатические влияния через а-адренорецепторы тормозят секрецию инсулина.
Стимулируют продукцию глюкозы (повышают ее уровень в крови) симпатические влияния и ряд контринсулярных гормонов: глюкагон в печени, адреналин через (32-адренорецепторы и ↑цАМФ в скелетных миоцитах и сердце в результате гликогенолиза, глюкокортикоиды (в результате гликогенолиза), глюкокортикоиды, глюкагон (в результате глюконеогенеза), СТГ (активирует секрецию глюкаго-на), тироксин (стимулирует глюкозо-6-фосфатазу).
Характеристика обмена жиров. Незаменимые жирные кислоты. Роль жиров в энергообеспечении организма. Регуляция жирового обмена.
Обмен липидов (составляют 10—20 % массы тела). Необходимо 100г жиров в сутки.
Жиры: триглицериды, фосфолипиды, холестерины, сфинголипиды и т.д.
Функции липидов:
1) пластическая — важный компонент клеточных мембран;
2) энергетическая — при окислении липиды дают максимальное образование энергии (1 г жиров — 9 ккал);
3) механическая — фиксация положения органов, уменьшение травмирования органов;
4) терморегуляторная — теплоизолирующие свойства подкожной клетчатки, теплопродукция в бурой жировой ткани;
5) используются для синтеза биологически активных веществ (простагландинов, простациклинов, тромбоксанов, лейкотриенов, стероидных гормонов).
Незаменимые жирные кислоты: холевая, фезоксихолевая, архидоновая кислота.
Жиры обладают наибольшей энергетической ценностью: 1 г жиров эквивалентен по крайней мере 9 ккал
Регуляция обмена липидов.
Нервная.
Симпатические влияния усиливают распад (норадреналин, β-адренорецепторы, ↑цАМФ) и тормозят синтез (α-адренорецепторы, ↓цАМФ) триглицеридов в жировой ткани.
Парасимпатические влияния способствуют отложению жира (ацетилхолин, М-холинорецепторы, ↓цАМФ).
Гуморальная.
•Активируют мобилизацию жира и его окисление адреналин (3-адренорецепторы, ТцАМФ) и гипогликемия (через торможение секреции инсулина); другие гормоны (СТГ, ТТГ, АКТГ, глюкагон, вазопрессин) у человека оказывают очень слабое действие.
•Тормозят мобилизацию жира и активируют его синтез инсулин (особенно в жировой ткани), глюкокортикоиды (депонирование жира в области лица и шеи), адреналин (α2-адренорецепторы, ↓цАМФ)
Обмен воды и минеральных солей в организме. Регуляция водного и минерального обменов.
Обмен воды
Человеку нужно в сутки 2-2,5 л воды. Это количество воды поступает из следующих источников:
Вода при питье(1л), в пище(1л), вода, образующаяся в организме при обмене белков, жиров и углеводов (300-350 мл). Основные органы, удаляющие воду из организма, - почки(1,2-1,5 л воды в составе мочи), потовые железы (500-700 мл), легкие (350мл) и кишечник(100-150мл).
Для нормальной деятельности организма важно, чтобы поступление воды полностью покрывало ее расход. Отношение количества потребленной воды к количеству выделенной составляет водный баланс.
Регуляция:
При недостатке воды в организме повышается осмотическое давление тканевой жидкости. Это приводит к раздражению расположенных в тканях особых рецепторов - осморецепторов. Импульсы от них по специальным нервам направляются в головной мозг к центру регуляции водно-солевого обмена. Оттуда возбуждение направляется к железе внутренней секреции - гипофизу, который выделяет в кровь специальный гормон, вызывающий задержку мочеотделения. Уменьшение выделения воды с мочой восстанавливает нарушенное равновесие.
Рефлекс начинается нервным путем с осморецепторов, а затем включается гуморальный механизм - поступление в кровь специального гормона.
Обмен солей
С наличием минеральных веществ связано явление возбудимости - одного из основных свойств живого. Рост и развитие костей, нервных элементов, мышц; они определяют реакцию крови (рН), способствуют нормальной деятельности сердца и нервной системы, используются для образования гемоглобина (железо), соляной кислоты желудочного сока (хлор).
Минеральные соли создают столь необходимое для жизнедеятельности клеток определенное осмотическое давление.
Макроэлементы:
Калий.
Функции К+: 1) главный внутриклеточный катион, определяющий осмотическое давление в клетках; 2) его диффузия из клетки является главным механизмом формирования мембранного потенциала и фазы репо-ляризации в потенциалах возбудимых клеток; 3) влияет на сосудистый тонус.
- Содержание К+ (в ммолях) во всем организме 3700— 4600 (~ 175 г), в скелетных мышцах ~ 2800, коже — 360—500, костях — 218, печень — 135—200, в внеклеточной жидкости ~ 60. Основным депо К+ являются скелетные мышцы.
- Потребность К+ в сутки 90—100 ммоль (~ 4,5 г), всасывается в кишечнике ~ 90 % К+ пищи.
- Концентрация К+ в межклеточной жидкости в основном определяется двумя факторами:
1) рН внеклеточной жидкости: уменьшение (увеличение) рН на 0,1 приводит к увеличению (уменьшению) К+ в крови нa 0,6 ммоль/л; 2) реабсорбцией и секрецией К+ в почечных канальцах.
- Все пищеварительные соки, молоко и пот содержат К+ в большей концентрации, чем плазма крови.
- Выделение К+ из организма осуществляется главным образом с мочой почками (80—90 ммоль/сут), калом (~ 10 ммоль/сут).
- Для баланса К+ в организме необходимо, чтобы его количество, выделенное с мочой, было равно количеству К+, всосавшемуся в кишечнике.
- Основным регулятором обмена К+ в организме является альдостерон.
Натрий.
- Функции Na+: 1) главный катион внеклеточных жидкостей, это определяет его роль в формировании осмотического равновесия этих жидкостей и обмена воды между клеточным и внеклеточным пространствами; 2) участвует в формировании фазы деполяризации ПД и других биопотенциалов; 3) участвует в регуляции сосудистого тонуса, повышает его.
- Баланс Na+. Организм человека содержит ~ 4200 ммоль (~ 95 г) Na+: во внеклеточных жидкостях -2100 (50%), в костях находится ~ 1800 (42 %), внутри клеток 260 (8 %) Na+.
- Поступает Na+ в организм в количестве 3—6 г в сутки; поступление Na+ меньше 1 г в сутки приводит к его отрицательному балансу с развитием гипонатриемии.
- Наибольшая часть поступившего в организм Na+ находится в коже и подкожной клетчатке, мышцах, хрящах; эти ткани рассматриваются как депо Na+.
- Почки являются главным органом, определяющим натриевое равновесие в организме, при этом главным регулятором является альдостерон.
- Na+ выделяется из организма с мочой (~ 90 %), потом и калом.
Кальций.
-Функции Са2+: 1) пластическая — входит в состав костей (10 % массы органа), зубов (эмаль — 34 %, дентин — 24 %); 2) влияет на возбудимость нервной и мышечной тканей; 3) необходим для свертывания крови; 4) регулирует проницаемость клеточной мембраны; 5) запускает мышечное сокращение, секрецию ферментов, гормонов и медиаторов; 6) является посредником при реализации эффектов многих гормонов; 7) влияет на репликацию ДНК и клеточное деление.
-Содержимое Са2+ в организме значительно превышает количество всех других минеральных веществ и равно примерно25 молей (1,1 кг), 99 % этого количества приходится на долю костей и зубов. В крови содержится примерно 2,5 ммоль/л Са2+, половина которого находится в ионизированном виде, другая половина — в комплексе с альбуминами.
- Потребность в Са2+ взрослого человека равна 0,5— 0,8 г/сут (~ 8 мг/кг массы), у детей, беременных и кормящих матерей она в 2—4 раза больше. Лучшими источниками Са2+ являются молоко, сыры, творог, фасоль, зеленый лук. Ухудшает всасывание Са2+ из кишечника большое содержание в пищи жиров, фосфатов и щавелевой кислоты, улучшают всасывание белки пищи, лактоза.
- Регулируют уровень Са2+ в крови паратгормон и 1,25-(OH)2-D3 (они повышают уровень Са2+ в результате мобилизации его из костей, увеличения реабсорбции в почках и всасывания в кишечнике), а также кальцитонин (снижает уровень Са2+ в результате перехода его в кости, выделения с мочой и торможения всасывания в кишечнике).
- Выделение Са2+ из организма происходит как с мочой (~ 30 %, 0,25 г/сут), так и через кишечник.
Хлор.
- Функции Cl-: 1) участие в регуляции водно-солевого обмена и осмотического давления во внеклеточной и клеточной жидкостях; 2) участие в формировании биопотенциалов клеток; 3) входит в состав соляной кислоты желудочного сока.
- Содержание Сl- в организме около 3 молей (110 г), суточная потребность около 5 г, он поступает в организм в основном за счет NaCl пищи, наибольшее количество Сl- (до 60 % всего Сl-) находится в коже и подкожной клетчатке.
- Выделение Сl- из организма происходит главным образом с мочой (около 140 ммоль/сут), а также с потом.
Микроэлементы (железо, йод, медь, цинк, кобальт, молибден, селен, хром, фтор, кремний и др.). Их функции связаны с тем, что они входят в состав многих ферментов: (напр., в оксидазах — медь, молибден, в глютатионпероксидазе — селен, в лактатдегидрогеназе — цинк); гормонов (напр., в Т3, Т4 — йод), гемоглобина и цитохромов — железо; витаминов (напр., В12 — кобальт).
Витамины, их характеристика, симптомы гиповитаминозов.
Витамины не являются энергетически богатыми соединениями, входят в состав разных ферменто или коафакторами ферментов.
Водорастворимые: входят в состав ферментов(софактор ферментов)
1)Витамины группы В
В1(тиамин): участвует во всех видах обмена, нормализует работу сердца, повышает секрецию и моторику желудка. При недостатке:бери-бери, полиневрит(разрушение миелиновой оболочки и развитие паралича) Находится в семенной кожуре риса, продуктах животного происхождения (в печени).
В2(рибофлавин): входит в состав ферментов. При недостатке у животных дерматиды, рвота, диарея; у человека коньюктивит, катаракта, головные боли, депрессии, мышечная слабость, миостения.
В3(никотиновая кислота): При недостатке: пеллагра, дерматит (повыш чувствительность кожи к солнцу), деменции (нарушение психических функций, галлюцинации, диарея)
В5(пантотеновая кислота): входит в состав ацетилкофермента а) При гиповитаминозе нарушается обмен белков, углеводов. Содержится во всех продуктах.
В6(пиридоксин): кофактор многих ферментов. При гиповитаминозе:дерматит, нарушение роста, дистрофия печени, у детей – развитие приступа эпилепсии.
В9(фолиевая кислота):получают из листа. Входит в всостав многих коферментов для процессов репликации ДНК. При недостатке – нарушение роста, анемия.
В12(цианкобаламин): для процесса эритропоэза. При недостатке: анемия, развитие демиелинизации. Для всасывания – фактор Касла в желудке, если его нет – анемия.
Н(биотин):синтезируется в печени, почках. При недостатке: специфический дерматит, увелечение секреции сальных желез(себорея), нарушение функционирования волосяных сумочек, депрессия, астения.
С(аскорбиновая кислота):является антиоксидантом. Недостаток: цинга (проявляется в кровотечении десен, воспалении зубов.) Содержится в продуктах ратсительного происхождения: цитрус, шиповник, фрукты.
Жирорастворимые витамины:
А(ретинол): входит в состав фотопигмента родопсина. Влияет на функцию зрения, обеспечивает адаптацию к свету, необходим для роста и дифференцировки эпит, нервной и костной ткани. Недостаток: нарушение зрения(куриная слепота), нарушение кожных покровов, роговица глаза становится прозрачной. Гипер:атрофия семенных канальцев. Находится в продуктах животного и растительного происхождения, каротиноиды в моркови.
D: D2(кальциферол), D3(холикальциферол), D4 (дигидрокальциферол). Недостаток: рахит (особенно у детей), остеомаляция(у взрослых), нарушение всасывания жиров. Образуется в почках. D3D4 под УФ
Е(токоферол): Нет ферментной системы антиоксидантного звена. Играет роль в репродукционных функциях, участвуетв анаболизме, необходим для мышечного развития, играет роль в митотическом делении клетки. Нарушение: анемия.
К(филлохитон): необходим для процесса свертывания крови, является кофактором для образования 4, 7, 9, 10 факторов свертывания крови, частично синтез микрофлоры кишечника. Находится в растительной пище.
Функциональная система поддержания оптимальной температуры организма Механизмы теплопродукции и теплоотдачи. Регуляция изотермии. Особенности терморегуляции в условиях гипо- и гипертермии.
Система поддержания оптимальной температуры организма
Химическая терморегуляция осущ путем усиления или ослабления образования тепла организмом, т.е. путем усиления или ослабления интенсивности его обмена веществ. Физическая терморегуляция осуществляется путем изменения интенсивности отдачи тепла телом.
Механизмы теплопродукции и теплоотдачи.
1)Теплопродукция
Сократительный термогенез – момент мышечного сокращения вызывает образование тепла. Несократительный термогенез – ионные насоса Na+\K+ - АТФаза бурый жир у спяцих животных энергия преобразуется в тепло.
2) Теплоотдача
Перенос части тепла в окружающую среду во время дыхания., излучение (если температура тела выше температуры окружающей среды), испарение (выделение через потовые железы воды и минеральных солей) Для испарения 1 мл воды затрачивается 0,5 ккал.
Регуляция тепла осуществляется нервным и гуморальным путем. Центр в гипоталамусе, при этом передние ядра обеспечивают регуляцию теплоотдачи (при нарушении – гипертермия), задний гипоталамус – центр теплопродукции. Активность СНС приводит к нарушению обмена веществ. Передний гипоталамус ПСНС – потоотделение, жажда, регул солевого обмена, увеличение излучения. Гипоталамус – орган оценки температуры тела, отслеживает колебания, термостат, ответственен за ряд нарушений процессов терморегуляции: лихорадка(нарушение терморегуляции при заболевании), действие токсинов(пирогенны), нарушение взаимодействия между центрами теплопродукции-теплоотдачи. С помощью применения воды для регуляции температуры потоотделение усиливается, с потом выделяются соли => развитие нарушения деятельности сердца, нервной системы.
Особенности терморегуляции в условиях гипертермии.
На первой стадии перегревания организма теплоотдача испарением влаги с поверхности тела компенсирует тепловую нагрузку. На второй стадии теплоотдачи испарением больше или равна внешней тепловой нагрузке. Третья стадия перегревания характеризуется уменьшением теплоотдачи с помощью испарения влаги, которая становится меньше тепловой нагрузки.
Вначале отмечаются явления, успешно противостоящие неблагоприятным воздействиям на организм высоких температур: увеличение теплоотдачи испарением влаги, понижение теплопродукции. Когда же организм не справляется с тепловой нагрузкой, усиливаются окислительные процессы, накапливается дополнительное тепло в организме. Испарение влаги с поверхности тела в это время достигает своего предела — начинается профузное потоотделение. А на третьей стадии наступает декомпенсация со стороны сердечно-сосудистой системы, приводящая к тепловому удару, признаками которого являются покраснение кожи, головокружение, головная боль, падение сердечной деятельности. При этом нарушается ритм дыхания, возникают тошнота и рвота вплоть до потери сознания.
Особенности терморегуляции в условиях гипотермии.
В начале охлаждения происходит некоторое повышение температуры тела вследствие перераспределения крови в организме: при сужении периферических кожных и подкожных кровеносных сосудов, которое в это время возникает, значительное количество крови отливает к внутренним органам, сосуды которых расширяются и в них увеличивается кровоток.
При длительном воздействии на организм холода включаются дополнительные механизмы, направленные, с одной стороны, на уменьшение теплоотдачи, а с другой — на повышение процессов теплопродукции. При этом усиливается деятельность сердца, появляется одышка, повышается мышечный тонус, возрастает мышечная активность, возникают озноб и дрожь. Все это способствует повышению обмена веществ в организме.
Продолжительное охлаждение вызывает увеличение секреции тиреотропного гормона передней доли гипофиза и повышение активности щитовидной железы. Гормон щитовидной железы — тироксин способствует стимуляции обмена веществ. Повышению обмена веществ в организме способствует и гормон надпочечников — адреналин. этот гормон усиливает скорость кровотока внутри организма. В результате предотвращается избыточное выделение тепла.
Дальнейшее охлаждение организма, при котором аксиллярная температура падает ниже 35°, приводит к глубокой гипотермии, которая сопровождается падением скорости обменных процессов, снижением газового обмена, падением кровяного давления, урежением сердечных сокращений и дыхания и т. д.: происходит дальнейшее падение температуры тела.
Функциональная система выделения, органы выделения. Функции почек. Нефрон как структурно-функциональная единица почки. Типы нефроиов. Особенности кровоснабжения почек.
Физиологическая система выделения — совокупность органов, взаимосвязанная деятельность которых обеспечивает постоянство ионного состава, осмоляльности, рН, объема жидкости сосудистого, интерстициального и внутриклеточного секторов, концентрации конечных продуктов обмена во внутренней среде организма.
Органы выделения: система дыхания, слюнные железы, печень, желудок, кишечник, кожа, сальные железы.
Функции почек:
1)Мочеобразование и мочевыделение
- обеспечивает осморегуляцию(Na+, K+, Ca2+, глюкоза)
- волюморегуляция(H2O) – поддержание объема циркулирующей крови, водный баланс организма
- экскреторная функция (мочевина, мочевая кислота, аммиак)
- регуляция артериального давления
- рН(Н+,НСО3-,SO42-)
2)метаболистические процессы ( в почках может происходить образование глюкозы – глюконеогенез – в условиях голодания.
3)гидролиз белков-олигопептидов могут гидролизоваться до аминокислот.
4)инкреторная функция (железа внутренней секреции, синтез биологически активных веществ) синтез эритропоэтинов, синтез ренина действие на ангиотензиноген образование ангиотензин – важный вазотрансгибитор.
5)синтез активной формы витамина D3 – регуляция обмена кальция, синтез калликреин брадикинин. (регуляция тонуса кр.сосудов
- особенности кровоснабжения 1,2 л\мин 42 мл\г
- с потреблением О2 =
Нефрон как структурно-функциональная единица почки. (~ 1 млн в почке).
Компоненты нефрона и их функция.
- Почечное тельце: сосудистый клубочек (50—100 капилляров) и капсула — осуществляет функцию фильтрации.
- Почечные канальцы (длина 35—50 мм) осуществляют функции реабсорбции, секреции, осмоконцентрирования и разведения.
- Проксимальный извитой каналец (25 % длины), щеточная кайма.
- Петля Генле (50 % длины): нисходящая часть (проксимальный прямой каналец и тонкий нисходящий каналец) и восходящая часть (тонкий и толстый канальцы).
- Дистальный извитой каналец (15 % длины). Соединительный каналец.
- Собирательная трубочка (в функциональном плане относят к нефрону, 10 % длины). Имеет светлые клетки, обеспечивающие реабсорбцию воды и темные клетки, которые секретируют Н+, закисляя мочу.
- Юкстагломерулярпый аппарат {ЮГА) — инкреторная функция.
- Эпителиоидные (зернистые) клетки приносящей и частично выносящей артериолы (секреция ренина, эритропоэтина).
- Клетки плотного пятна дистального извитого канальца (осморецепторы, реагирующие на осмотическое давление канальцевой жидкости).
- Агранулярные клетки Гурмагтига передают влияние с клеток плотного пятна на эпителиоидные клетки и являются резервом по секреции ренина.
- Мезапгиальные клетки благодаря сокращению регулируют кровоток в клубочке, обладают фагоцитарной активностью, образуют матрикс мезангия, секретируют про-стагландины и др. Основные виды нефронов.
- Корковые нефроны имеют короткие канальцы, в них преобладают функции фильтрации, реабсорбции, секреции, менее выражена осмоконцентрация мочи.
- Юкстамедуллярные нефроны (их в 4 раза меньше) имеют длинную петлю Генле, в них резко выражена функция концентрирования и разведения мочи.
Кровообращение в почке.
Особенности сосудистой сети почек.
Двойная сеть капилляров:
- приносящие артериолы дают первую капиллярную клубочковую сеть с высоким давлением крови (~ 60 мм рт. ст.), что способствует фильтрации;
- выносящая из клубочка артериола распадается на вторую капиллярную сеть вокруг канальцев с низким давлением крови (~ 10 мм рт. ст.), что способствует реабсорбции.
От выносящих артериол отходят также прямые сосуды, которые повторяют ход петель Генле в мозговом веществе; кровоток по прямым сосудам составляет ~ 2 % почечного кровотока; эти сосуды играют важную роль в противоточном осмоконцентрирующем механизме.
Характеристика кровотока в почках.
Высокий уровень кровотока: 1000—1200 мл/мин на 1,73 м2 поверхности тела или 450 мл/100 г ткани почки в 1 мин, что составляет 20—25 % сердечного выброса.
Высокий уровень потребляемого кислорода (6—10%, ~ 18 мл/мин) при низкой величине артериовенозной разницы — 14 мл 02/л крови.
Ауторегуляция почечного кровотока в диапазоне 80— 180 мм рт. ст. системного артериального давления.
Два круга кровообращения в почках.
- Кровоток в корковом веществе (составляет 90 %, осуществляется приносящими артериолами корковых нефронов, отходящими от дистальных участков междольковых артерий).
- Кровоток в мозговом веществе (составляет 10 %, осуществляется по приносящим артериолам юкстамедуллярных нефронов, отходящих от проксимальных участков междольковых артерий, а также по прямым артериолам, отходящими непосредственно от дуговых артерий).
- Создается возможность (она реализуется при централизации кровотока) путем сужения междольковой артерии между приносящими артериолами корковых и юкстамедуллярных нефронов выключить кровоток в корковом веществе и значительно уменьшить фильтрацию.
Клубочковая фильтрация и ее механизмы. Состав и свойства первичной мочи. Измерение скорости клубочковой фильтрации (клиренс).
Фильтрация протекает в капсуле и заключается в образовании первичной мочи. Как предполагается, первичная моча фильтруется из капилляров мальпигиева клубочка в полость капсулы. Для того чтобы была возможна фильтрация, необходима значительная разность давления в сосудах и капсуле. Сила эффективного фильтрационного давления = Рарт – Ронкот.давл. – Рультрафильтр= 50-25-15=10
Фильтрации подвергается плазма (нефильтрующиеся белки молекулярная масса > 70 кД, форменные элементы крови, наличие белка – протенурия), ионы солей, глюкоза, витамины, продукты метаболизма белков, мочевина, мочевая кислота. Объем – 1,2 л\мин, за сутки – 170-180 л первичной мочи.
Состав:Первичная моча содержит необходимую организму воду и растворенные в ней вещества, большинство из которых представляют биологическую ценность, например аминокислоты, углеводы, соли, витамины, продукты обмена и др. Лишь некоторые из веществ, растворенных в первичной моче, безусловно, требуют выведения из организма. К ним относятся мочевина, мочевые кислоты, креатинин, сульфаты и др.
Объем фильтрации можно определить с помощью инулина (не абсорбируется, не метаболизируется) клиренс инсулина – показатель, помогающий оценить объем.
Клирекс = Vи * [Hи]\ [Uг] – объем фильтр. в нейронах почек.
Реабсорбция в проксимальном канальце нефрона, особенности реабсорбции аминокислот и глюкозы. Осмотическое разведение и концентрация мочи; поворотно-противоточная система нефрона. Канальцевая секреция и ее механизмы.
Реабсорбция (обратное всасывание) протекает в канальцах нефронов, в большей степени вода по осмотическому градиенту в проксимальном канальце(60-70%), в петле Генли(20%), в дистальном канальце(10-20%). Клиатин – не реабсорбируется.
Реабсорбция в проксимальном канальце нефрона.
Участвует Na+\К+ АТФаза, обеспечивает реабсорбцию натрия - Na+\H+. Реабсорбируется 50% мочевины, витаминов, но часть витаминов теряется, глюкозы, которая (на апекальной мембране) транспортируется вместе Na+ по градиенту концентрации, на базальной мембране облегченная диффузия. Na и глюкоза относятся к пороговым веществам. Если концентрация глюкозы выше 8-10 ммоль\л, то глюкоза остается в моче.(при сахарном диабете) - глюкозурия; ангиопептиды - короткие белки, причем они будут подвергаться гидролизу в клетках канальцев (АМК). Важная роль в транспорте натрия играют протоны водорода:регулирует рН И реабсорбирует натрий и воду. Гиподинапия - повышение нагрузки на почки при употреблении диоретиков, что блокирует карбоангидразу.
Реабсорбция аминокислот (95—99 % профильтрованных).
Топография: проксимальные канальцы.
Механизм: на апикальной мембране происходит вторичный активный транспорт с использованием градиента Na+ и пяти типов переносчиков (для кислых, основных, нейтральных аминокислот, иминокислот, всех остальных аминокислот); на базальной мембране — облегченная диффузия.
Осмотическое разведение и концентрация мочи; поворотно-противоточная система нефрона.
В зависимости от состояния водного баланса организма почки могут выделять гипотоническую, очень разведенную или осмотически концентрированную мочу. В этом процессе участвуют все отделы канальцев и сосуды мозгового вещества почки, функционирующие как противоточная поворотная множительная система.
система поворотно-противоточная - физиологический механизм реабсорбции воды и ионов натрия в петле нефрона, основанный на различной проницаемости для них стенок восходящего и нисходящего колен петли и заключающийся в том, что реабсорбция воды в восходящем колене способствует реабсорбции натрия в нисходящем и наоборот.
Осмоконцентрирование мочи (противоточно-множительный механизм).
- Нарастающий градиент осмотического давления в межклеточной жидкости от границы коры и мозгового слоя(~ 300 мосм/кг) до вершины пирамид (~ 1200 мосм/кг) является основой концентрирующего механизма и создается:активной реабсорбцией NaCl в толстом сегменте восходящей части петли; реабсорбцией мочевины из нижних отделов собирательных трубочек (место высокой проницаемости для мочевины); задержкой этих веществ в интерстиций с помощью сосудистой (прямые сосуды) и канальцевой (восходящая часть петли — собирательная трубочка) противоточных систем.
- В нисходящий тонкий отдел петли Генле, имеющий высокую проницаемость для воды и низкую для электролитов: поступает моча из проксимального канальца, изотоничная плазме (~ 300 мосм/кг); при продвижении фильтрата вода выходит в интерстиций против осмотического давления, а электролиты и мочевина остаются в канальце; в результате увеличивается осмотическое давление фильтрата в нисходящей части петли от 300 до 1200 мосм/кг в области вершины петли;при этом объем фильтрата уменьшается на 18 % (на ~ 35 л). Деуретики блокируют реабсорбцию воды, натрия; петлевые деорутики – выводят воду с мочой из организма.
- В восходящую часть петли Генле имеется плотное пятно – рецепторный аппарат, который решает в какой степени осуществляется реабсорбция на последнем 3 канале. Благодаря ему нефроны очень быстро оценивают потребность в воде и NaCl,) имеющую высокую проницаемость для ионов и низкую для воды: поступает гиперосмотичный фильтрат;здесь происходит активная реабсорбция Na+, К+, Сl-; в результате в дистальный каналец поступает гипотоничная жидкость; здесь продолжается реабсорбция Na+, Сl- и воды, поэтому осмотическое давление существенно не меняется, а объем жидкости уменьшается.
- В собирательные трубки поступает гипо- или изотоничная моча:
здесь происходит осмотическое концентрирование мочи (максимально до 1400 мосм/кг), так как вода реабсорбируется не только следуя за Na+ и Сl-, но и против высокого осмотического давления интерстиция; реабсорбция воды в собирательных трубочках контролируется АДГ.
Осмотическое разведение мочи (водный диурез) возникает при водной нагрузке (а также нарушении секреции или эффекта действия АДГ).
Снижение секреции АДГ при водной нагрузке приводит к блокаде реабсорбции воды в собирательных трубочках при сохраняющейся реабсорбции Na+ и Сl- в нефроне.
Максимальная осмоляльность в интерстиций снижается при этом до 500 мосм/кг.
Максимальный водный диурез равен от 12 л/сут (контроль АДГ при нормальном объеме фильтрата) до 30 л/сут при увеличении фильтрации в результате повышенного приема жидкости.
Минимальное осмотическое давление мочи снижается до 40 мосм/кг.
Методы исследования: 1) проба с разведением (водная нагрузка — 1 л): снижается относительная плотность мочи до 1,002, осмотическое давление до 50 мосм/кг; 2) определение осмотического индекса (UM0CM / Рмосм): снижение до 0,2 (норма ~ 2).
Осмотический диурез – выведение мочи с водой. Вода может выводиться, т.к. может быть высокая концентрация натрия, мочевиды, глюкозы.
Полиурия – осмотический диурез.
Мочевина играет роль в регуляции диуреза.
Гипертоницеский диурез – повышение АД, что способствует повышению давления, выделению большего кол-ва мочи.
Канальцевая секреция (процесс перехода веществ из крови через канальцы в мочу, а также из клеток канальцев в мочу или в кровь).
Топография секреции:
- в проксимальных канальцах: Н+, NH3, мочевая кислота, холин, серотонин, пенициллин, ПАГ, фенолрот и др.;
- в тонкой части петли: мочевина;
- в дистальных канальцах и собирательных трубочках: Н+, NH3 и К+ при избыточном поступлении с пищей.
Секреция К+ (при избыточном поступлении с пищей).
Топография: дистальные канальцы и собирательные трубочки.
Механизм: К+, Na+-Hacoc на базальной мембране, диффузия через каналы на апикальной мембране, контролируется альдостероном; часть К+ секретируется по межклеточным щелям через плотные контакты.
Секреция Н+ из клетки канальца в мочу (~ 75 ммоль/сут).
Топография: проксимальные, дистальные канальцы, собирательные трубочки.
Механизм: Н+ освобождается при диссоциации образующейся Н2С03; на апикальной мембране клеток проксимальных канальцев выделяется в просвет канальцев путем ионообмена (антипорта) с Na+, в собирательных трубочках — с участием Н+-насоса.
Секреция NH3 из клеток канальцев в мочу (~ 50 ммоль/сут).
Топография: проксимальные и дистальные канальцы, собирательные трубки.
Механизм: NH3 образуется в клетке в реакциях дезамидирования глутамина и дезаминирования глутамата, на апикальной мембране происходит диффузия NH3 в просвет канальца.
Принцип строения анализаторов (сенсорных систем). Классификация и характеристика рецепторов. Принципы организации и основные функции сенсорных систем (обнаружение, различение, преобразование, кодирование). Адаптация сенсорных систем.
Анализаторы – системы, обеспечивающие взаимодействие организма с окружающей средой.
Принцип строения:
1) многослойность, то есть наличие нескольких слоев нервных клеток, первый из которых связан с рецепторами, а последний – с нейронами моторных областей коры большого мозга. Это свойство дает возможность специализировать нейронные слои на переработке разных видов сенсорной информации, что позволяет организму быстро реагировать на простые сигналы, анализируемые уже на первых уровнях сенсорной системы. Создаются также условия для избирательного регулирования свойств нейронных слоев путем восходящих влияний из других отделов мозга;
2) многоканальность сенсорной системы, то есть наличие в каждом слое множества (от десятков тысяч до миллионов) нервных клеток, связанных с множеством клеток следующего слоя;
3) разное число элементов в соседних слоях, что формирует «сенсорные воронки»;
4) дифференциация сенсорной системы по вертикали и по горизонтали. Дифференциация по вертикали заключается в образовании отделов, каждый из которых состоит из нескольких нейронных слоев. Дифференциация по горизонтали заключается в различных свойствах рецепторов, нейронов и связей между ними в пределах каждого из слоев.
Классификафия рецепторов:
1)по структуре рецепторов: свободные нервные окончания (болевые, температурные, некоторые механорецепторы), инкапсулированные нервные окончания (тельца Пачини, Мейсснера и Руффини, колбы Краузе, мышечные веретёна, сухожильные рецепторы Гольджи), специализированный нейрон (напр., фоторецепторы, центральные хеморецепторы), эпителиальная волосковая клетка (напр., слуховые и вестибулярные рецепторы).
2)По расположению в организме: экстерорецепторы и интерорецепторы, центральные и периферические рецепторы.
3)В зависимости от вида раздражителя: механо-, хемо-, термо-, фоторецепторы, ноцицепторы.
4)По психофизиологическому критерию: зрительные, слуховые, вкусовые, обонятельные, тактильные.
5)По степени специфичности: моно- и полимодальные рецепторы.
6)По скорости адаптации: быстро- и медленно адаптирующиеся, смешанные.
7)По электрофизиологическому критерию: первичные рецепторы (рецепторный потенциал и потенциал действия возникают в одной клетке) и вторичные рецепторы — РП возникает в волосковой клетке или фоторецепторе, а ПД — в нейроне (напр., зрительные, слуховые, вестибулярные).
Функции:
1)Обнаружение сигналов.
- Минимальный порог возбуждения — сила раздражителя, вызывающая вероятность восприятия в 50— 75 % случаев его действия.
- Максимальный порог возбуждения — сила раздражителя, вызывающая болевую реакцию.
2)Различение сигнала (дифференциальные пороги).
- Порог интенсивности — минимальное изменение силы раздражителя, которое позволяет ощущать его как новый раздражитель. Напр., для обонятельного анализатора дифференциальный порог интенсивности равен 30%, для вкусового ~ 10 %, тактильного ~ 3 %, зрительного ~ 1,5 %, слухового ~ 1 %.
- Пространственные пороги — минимальное расстояние между двумя раздражителями, при котором они ощущаются раздельно. Напр., на кончике языка ~ 1,1 мм, пальца ~ 2,5 мм, на плече ~ 68 мм.
- Временные пороги — минимальное время между двумя раздражителями, при котором они воспринимаются раздельно (напр., в зрительном анализаторе — 0,04 с для палочек).
3)Преобразование
4)Кодирование информации в рецепторном отделе.
- Кодирование силы раздражителя амплитудой рецепторного потенциала.
- Кодирование продолжительности действия раздражителя продолжительностью рецепторного потенциала (в медленно адаптирующихся рецепторах).
- Кодирование "начало-конец" раздражения (характерно для быстро адаптирующихся рецепторов).
- Кодирование направления движения стимула (напр., в вестибулярных рецепторах).
Адаптация анализаторов — это процесс приспособления к оптимальному восприятию раздражителей среды в связи с потребностями организма.
1)Адаптационные процессы на дорецепторном уровне (напр., рефлекторная реакция зрачка и хрусталика, сокращение мышц среднего уха, поворот глаз и головы в сторону раздражителя).
2)Адаптационные процессы на рецепторном уровне.
•Снижение возбудимости рецепторов.
- Кальциевый механизм: повышение концентрации Са2+ в рецепторе активирует Са2+-зависимые К+-каналы, что препятствует фазе деполяризации и усиливает фазу реполяризации рецепторного потенциала.
- Фосфорилирование рецепторных молекул (напр., родопсина при действии света, обонятельных молекул при действии цАМФ-зависимой протеинкиназы А), которые утрачивают свойство активировать С8-белки и запускать каскад реакций, приводящий к формированию РП.
- Инактивация Ка+-каналов в зоне перехода рецепторного потенциала в потенциал действия.
•Повышение возбудимости рецепторов.
- Увеличение количества функционирующих рецепторных молекул: их активация (напр., молекул родопсина в темноте) и увеличение их синтеза.
- Увеличение температуры в области рецепторов (активация ионных каналов).
3) Адаптационные процессы на центральном уровне.
- Эфферентные тормозящие и возбуждающие влияния на рецепторы (их функциональная мобильность) и центры переключения проводникового отдела со стороны вышележащих отделов ЦНС.
- Влияния на рецепторы и центры сенсорной системы через эндокринную систему.
- Изменение возбудимости корковых сенсорных зон (роль ретикулярной формации, лимбической системы и охранительного торможения).
Могут адаптироваться. Это связано с чувствительностью рецепторов – величина непостоянная. Абсолютный порог чувствительности – минимальная сила раздражителя, вызывающая возбуждение. Порог различения практически всегда выше ранее действовавшего раздражения на определенную долю (закон Вебера). Закон Вебера-Фехнера ∆I\I=const. Прирост силы раздражителя по отношению к анализатору – величина постоянная.
Зрительный анализатор. Вспомогательные аппараты глаза. Строение оптического аппарата глаза. Аккомодация и ее механизмы. Аномалии рефракции глаза. Зрачковый рефлекс.
Зрительный анализатор – это сенсорная система, воспринимающая электромагнитные излучения в видимом диапазоне и формирующая световые ощущения (обеспечивает поступление в мозг около 90% информации).
Периферическая часть – оптический аппарат глаза + вспомогательные аппараты.
Проводниковая часть – 2ая пара черепных нервов.
Центральная часть – верхние холмики среднего мозга, латеральные коленчатые тела промежуточного мозга, зрительная кора.
Вспомогательные органы глаза
Веки , В области медиального угла, имеется слезное озеро. Сверху веко ограничивается бровью, поросшей короткими жесткими волосами. К верхнему веку подходит мышца, поднимающая верхнее веко. По краям век располагаются в 2-3 ряда ресницы.
Коньюктива – это тонкая соединительнотканная оболочка, в которой выделяют коньюктиву век, покрывающий изнутри веки, и коньюктиву глазного яблока. Пространство, расположенное спереди от глазного яблока и ограниченное коньюктивой – коньюктивный мешок. У медиального угла глаза расположено слезное мясцо.
Слезный аппарат глаза включает слезную железу и систему слезных путей.
Слеза оттекает в слезное озеро у медиального края глаза, по краям век имеются верхний и нижний слезные сосочки, в каждом есть отверстие – слезная точка, дающая начало слезному канальцу, которые впадают в слезный мешок. Нижний конец мешка переходит в носослезный проток, открывающийся в нижний носовой ход.
Движение глазных яблок осуществляется 6 глазодвигательных мышц: 4 прямые и 2 косые. Прямые поворачивают глазное яблоко в соотв. Направлении, косые вокруг сагиттальной оси: верхняя вниз и кнаружи, нижняя вверх и кнаружи.
Строение оптического аппарата глаза:
Глаз состоит из:
Сосудистой оболочки, сетчатки, ресничатого тела, линзы, радужки. Имеются циркулярные мышцы (ПСНС) – сужение зрачка) и радиальные (СИС – расширение). Атропин блокирует циркулярные мышцы.
Сетчатка глаза: фоторецепторы(палочки и колбочки) ,биполярные и ганглионарные нейроны. Палочки и колбочки прилежат к сосудистой оболочке, за ними идут биполярные, а за ними – ганглионарные нейроны, а от них – 2 пара черепных нервов.
Палочки состоят из:
- Наружный сегмент (содержит мембранные диски с фотопигментом родопсином – состоящего из белка опсина + ретиталь (А)),
- Связующий отдел (ресничка),
- Внутренний сегмент (содержит митохондрии),
- Область с нервными окончаниями.
Строение колбочки:
- наружный сегмент (содержит мембранные полудиски),
- связующий отдел (перетяжка),
- внутренний сегмент (содержит митохондрии),
- синаптическую область.
Аккомодация и ее механизмы. Аномалии рефракции глаза. Зрачковый рефлекс
Рефлекс аккомодации глаза — это приспособление глаз к четкому видению различно удаленных предметов. Основным компонентом его является изменение кривизны хрусталика.
Рефлекторная реакция хрусталика.
Кривизна хрусталика зависит от: 1) его эластичности; 2) сил, развиваемых цилиарными мышцами, сосудистой оболочкой и склерой, воздействующих па сумку хрусталика; 3) внутриглазного давления.
- Рефлекторный путь: рецепторы сетчатки (стимул — нечеткое изображение предмета) —> зрительные нервы и тракты —> непарное парасимпатическое ядро III нерва —> цилиарный ганглий, сокращение цилиарных мышц (ацетилхолин + М-холинорецепторы) —> ослабление натяжения связок сумки хрусталика увеличение кривизны хрусталика.
- Диапазон аккомодации 10—14 D, что позволяет фокусировать предметы, находящиеся на 7—10 см от глаза.
•Рефлекс аккомодации глаза включает в себя также рефлекторное сужение зрачка и конвергенцию глаз.
Рефлекс зрачка — сужение или расширение зрачка, регулирующие световой поток на сетчатку в зависимости от степени освещенности.
•Рецепторы сетчатки (увеличение освещенности) —> зрительные нервы и тракты —> парные парасимпатические ядра III нерва —> цилиарный ганглий —> сокращение сфинктера радужки (ацетилхолин + М-холинорецепторы) —> сужение зрачка.
•Рецепторы сетчатки (уменьшение освещенности) —» цилиоспинальный центр (С8—Th,) —> верхний шейный ганглий —> сокращение радиальной мышцы радужки (норадреналин + а-адренорецепторы) -> расширение зрачка.
•Диапазон изменения освещенности сетчатки в результате зрачкового рефлекса ~ 30 раз (диаметр зрачка 1,5—8,0 мм).
Аномалии рефракции глаза.
Миопия (фокусировка предметов перед сетчаткой, оптическую силу глаза уменьшают вогнутыми линзами).
Гиперметропия (фокусировка предметов за сетчаткой, оптическую силу глаза увеличивают выпуклыми линзами).
Астигматизм — неодинаковая кривизна хрусталика.
Характеристика зрительного анализатора: абсолютная и дифференциальная зрительная чувствительность, световая и темновая адаптация; острота зрения, поля зрения; бинокулярное зрение; цветовое зрение и его нарушения, теории цветоощущения
Абсолютная чувствительность зрения.
Чтобы возникло зрительное ощущение, свет должен обладать некоторой минимальной (пороговой) энергией. Минимальное количество квантов света, необходимое для возникновения ощущения света в темноте, колеблется от 8 до 47. Одна палочка может быть возбуждена всего 1 квантом света. Таким образом, чувствительность рецепторов сетчатки в наиболее благоприятных условиях световосприятия предельна. Одиночные палочки и колбочки сетчатки различаются по световой чувствительности незначительно. Однако количество фоторецепторов, посылающих сигналы на одну ганглиозную клетку, в центре и на периферии сетчатки различно. Количество колбочек в рецептивном поле в центре сетчатки примерно в 100 раз меньше количества палочек в рецептивном поле на периферии сетчатки. Соответственно и чувствительность палочковой системы в 100 раз выше, чем у колбочковой.
Зрительная адаптация
При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной системы к условиям яркой освещённости называется световой адаптацией. световая адаптация возникает в результате фосфорилирования родопсина ферментом опсикиназой, фосфородопсин не способен активировать Gj-белок;
Обратное явление (темновая адаптация) наблюдается, когда из светлого помещения человек переходит в почти не освещённое помещение. В первое время он почти ничего не видит из-за пониженной возбудимости фоторецепторов и зрительных нейронов. Постепенно начинают выявляться контуры предметов, а затем различаются и их детали, так как чувствительность фоторецепторов и зрительных нейронов в темноте постепенно повышается. Темповая адаптация возникает в результате ингибирования опсинкиназы белком орестином в присутствии Са2+, что приводит к восстановлению активности молекул родопсина. (в темноте мембрана палочек открыта для ионов натрия)
Повышение световой чувствительности во время пребывания в темноте происходит неравномерно: в первые 10 мин она увеличивается в десятки раз, а затем, в течение часа - в десятки тысяч раз. Важную роль в этом процессе играет восстановление зрительных пигментов. Так как в темноте чувствительны только палочки, слабо освещённый предмет виден лишь периферическим зрением. Существенную роль в адаптации, помимо зрительных пигментов, играет переключение связей между элементами сетчатки. В темноте площадь возбудительного центра рецептивного поля ганглиозной клетки увеличивается из-за ослабления кольцевого торможения, что приводит к увеличению световой чувствительности. Световая чувствительность глаза зависит и от влияний, идущих со стороны мозга. Освещение одного глаза понижает световую чувствительность неосвещённого глаза. Кроме того, на чувствительность к свету оказывают влияние также звуковые, обонятельные и вкусовые сигналы.
Дифференциальная чувствительность зрения
Если на освещённую поверхность с яркостью I падает добавочное освещение dI, то, согласно закону Вебера, человек заметит разницу в освещённости только если dI/I = K, где K константа, равная 0,01-0,015. Величину dI/I называют дифференциальным порогом световой чувствительности. Отношение dI/I при разных освещённостях постоянно и означает, что для восприятия разницы в освещённости двух поверхностей одна из них должна быть ярче другой на 1 - 1,5 %.
Острота зрения:
Остроту зрения определяет то наименьшее расстояние между двумя точками, которое глаз может различить. Способность различать расстояние между точками зависит от угла, под которым видны эти точки. Нормальный глаз различает две точки под углом в 60 секунд. Максимальной остротой зрения обладает желтое пятно. К периферии от него острота зрения много ниже.
Острота зрения измеряется при помощи специальных таблиц, которые состоят из нескольких рядов букв или незамкнутых окружностей ной величины. Против каждой строчки стоит число, означающее стояние в метрах, с которого нормальный глаз должен различить этой строчки.
Острота зрения, определенная по таблице, выражается обычно в относительных величинах, причем нормальная острота принимается за единицу.
Поле зрения:
Если фиксировать глазом какую-либо точку, то изображение падает на желтое пятно; в этом случае мы видим точку центральным зрением. Точки, изображения которых падают на остальные места сетчатки, видимы периферическим зрением.
Совокупность точек, одновременно видимых глазом при фиксации взгляда в одной точке, называется полем зрения.
Измерение границы поля периферического зрения производят прибором, называемым периметром. Границы поля зрения для бесцветных предметов составляют к кверху 60° и кнаружи 90°. Поля зрения обоих глаз у человека совпадают, что имеет большое значение при восприятии пространства. Поля зрения для различных цветов неодинаковы, больше зрения для бесцветных предметов. Для синего и желтого цветов значительно меньше, для красного — еще меньше, а для зеленого стирается только на 20—30—40°.
Бинокулярное восприятие пространства осуществляется путем:
оценки удаленности предметов на основе дивергенции и конвергенции глаз при их восприятии;
стереопсиса (стереовосприятия) — наличия небольших различий в изображениях предмета, проецируемых на сетчатки обоих глаз, что приводит к впечатлению объемности (полное совпадение дает восприятие плоского изображения; более сильное несовпадение дает двоение зрительного образа);
"физиологического двоения" как механизма оценки глубины: когда изображения фиксируемого объекта совпадают на сетчатках обоих глаз, остальные объекты дают несовпадающие проекции, но двоение их изображения не воспринимается как таковое, а служит сигналом удаленности этих объектов относительно предмета наблюдения.
Цветовое зрение и его нарушения, теории цветоощущения.
трихромазия — нормальное цветовосприятие.
Трехкомпонентная теория цветного зрения (Ломоносов; Гельмгольц) предполагает наличие 3 типов колбочек со зрительными пигментами, поглощающими различные длины волн света: синие, зеленые, красные колбочки.
Теория оппонентных цветов (Геринг).
Имеются 4 цвета, которые образуют две парные антагонистические (оппонентные) группы (синий — желтый, зеленый — красный).
Соответственно имеется 4 вида ганглиозных клеток, образующих систему нейронов оппонентных цветов. Ганглиозные нейроны имеют 2 типа рецепторных полей с антагонизмом центра и периферии (желто-синие и красно-зеленые).Другие цвета воспринимаются путем "смешения" основных цветов.
Полихроматическая теория(Хартридж)
Наличие в сетчатке 7 типов рецепторов.
Нарушения восприятия цвета:
Врожденные формы цветовой слепоты (старое название — дальтонизм) связаны с отсутствием генов, кодирующих разные виды опсина в колбочках .
Дихромазия (отсутствие восприятия одного цвета). Дейтеранопия — "зеленослепые".Протанопия — "краенослепые". Тританопия — "синеслепые".
Ахромазия — полная цветовая слепота (черно-белое восприятие).
Морфофункциональная организация сетчатки глаза. Строение фоторецепторов. Молекулярно-клеточные механизмы фоторецепции. Особенности организации зрительных проводящих путей (зрительных нервов). Зрительные подкорковые центры и их функции.
Морфофункциональная организация сетчатки глаза.
Сетчатка представляет собой внутреннюю сетчатку глаза, в которой расположены фоторецепторы – палочки и колбочки, воспринимающие световые лучи, и нервные клетки с их многочисленными отростками, составляющие нервный аппарат глаза. В каждой сетчатке человека находится 6-7 млн. колбочек и 110-125 млн. палочек, которые располагаются неравномерно. Центр сетчатки содержит лишь колбочки. Количество колбочек уменьшается к периферии, где расположены только палочки. Самый наружные слой сетчатки образован пигментным эпителим, содержащим пигмент – фусцин. Этот пигмент поглощает свет, препятствуя отражению и рассеиванию его, что способствует четкости зрительного восприятия. Сетчатка глаза состоит: фоторецепторы(палочки и колбочки) ,биполярные и ганглионарные нейроны. Палочки и колбочки прилежат к сосудистой оболочке, за ними идут биполярные, а за ними – ганглионарные нейроны, а от них – 2 пара черепных нервов.
Функции:
1)Пигментный эпителий: регенерация зрительного пурпура после его обесцвечивания, а также принимает активное участие в обновлении наружных сегментов зрительных клеток, в защите рецепторов от светового повреждения, а также в переносе к ним кислорода и питательных веществ.
2)Колбочки обеспечивают дневное и цветовое зрение; более светочувствительные палочки ответственны за сумеречное зрение.
Строение фоторецепторов:
Палочки состоят из:
- Наружный сегмент (содержит мембранные диски с фотопигментом родопсином – состоящего из белка опсина + ретиталь (А)),
- Связующий отдел (ресничка),
- Внутренний сегмент (содержит митохондрии),
- Область с нервными окончаниями.
Строение колбочки:
- наружный сегмент (содержит мембранные полудиски),
- связующий отдел (перетяжка),
- внутренний сегмент (содержит митохондрии),
- синаптическую область.
Молекулярно-клеточные механизмы фоторецепции
Приводит к появлению ощущения света:
Свет попадает на родопсин и происходит образование цистранс ретиналь, это активирует фосфодиэстеразу, она участвует в активации трансдуцина (особый белок), который вызывает уменьшение количества ГМФ, а это блокирует входящий ток натрия. Эти реакции протекают очень быстро. В темноте мембрана палочек открыта для ионов натрия. Гиперполяризация мембраны – не поступает натрий.
В колбочках фотопигмент йодопсин.
Особенности организации зрительных проводящих путей (зрительных нервов). Зрительные подкорковые центры и их функции.
Аксоны ганглиозных клеток образуют зрительный нерв (II пара, ~ 1,3 млн волокон).
Зрительные нервы впереди серого бугра гипоталамуса образуют частичный перекрест (хиазму), где нервные волокна, идущие от носовых половин обеих сетчаток, пересекаются и переходят на противоположную сторону, а волокна, идущие от височных половин каждой сетчатки, не пересекаются.
После перекреста волокна образуют зрительный тракт, основные проекции которого:
в латеральные коленчатые тела и подушку (далее в первичную зрительную кору);
в верхнее двухолмие и далее в глазодвигательные центры ствола — ядра III, IV, VI нервов (зрительный ориентировочный рефлекс, рефлекторные реакции зрачка и хрусталика);
в верхнее двухолмие и далее через подушку таламуса, в конечном счете в теменную ассоциативную кору (поле 7), регулирующие произвольные движения глаз в контрлатеральную сторону;
в супрахиазматическое ядро гипоталамуса (регуляция суточных ритмов).
Зрительные тракты заканчиваются у подкорковых зрительных центров, основными из которых являются наружные коленчатые тела, передние бугры четверохолмия и подушки зрительных бугров.
Обработка сигналов в верхнем двухолмии, здесь имеются:
❖Нейроны, отвечающие на движения зрительного стимула в определенном направлении.
❖Нейроны, центрирующие изображение движущегося предмета в зоне центральной ямки и управляющие рефлекторным поворотом головы и глаз при внезапном появлении на периферии поля зрения движущегося объекта (зрительный ориентировочный рефлекс). Обработка сигналов в латеральных коленчатых телах.
Здесь осуществляется:
1) оценка степени освещенности; 2) оценка контрастности;
3) оценка цветовой характеристики стимула. Имеется топографическая проекция сетчатки: проекция центральной ямки занимает наибольшую часть.
56. Слуховой анализатор и его характеристика (звуковая чувствительность, громкость, бинауральный слух). Строение и функции наружного и среднего уха. Строение и функции внутреннего уха, кортиев орган слуховой улитки. Механизмы слуховой рецепции, электрические эффекты в улитке. Слуховые подкорковые центры и их функции.
Слуховой анализатор – сенсорная система, воспринимающая звуковые колебания внешней и внутренней среды (частотой от 16-20 Гц до 16-20 кГц) и формирующая звуковые ощущения.
Наибольшая чувствительность слуха в диапазоне 1— 4 кГц, в котором абсолютный порог интенсивности звука минимален. Резко снижена чувствительность при низких частотах (напр., при 100 Гц на 60 дБ). Звук, давлением свыше 140 дБ (107 выше порогового), вызывает боль и разрушение стереоцилий рецепторов.
Дифференциальные пороги слуховой чувствительности: (при частоте 0,5—2 кГц и интенсивности звука 40—50 дБ) порог интенсивности звука равен 0,5—1,0 дБ, порог частоты 1 %, длительности < 10 %, угла расположения источника звука 1—3°.
Пространственное восприятие звуковых раздражителей (бинауральный слух) реализуется на всех уровнях конвергенции (верхняя олива, нижнее двухолмие, слуховая кора) и создается:
при действии низких частот (до 1,3 кГц) различением времени поступления звука в правое и левое ухо (улавливается разница в 30 мкс);
при действии более высоких частот — различением интенсивности звука, поступающего в правое и левое ухо.