

1. Понятие о функциональных системах организма. Результат как системообразующий фактор. Принципы теории функциональных систем: саморегуляции, изоморфизма, мультипараметричности, иерархичности, взаимосодействия, системогенеза.
Теория функциональных систем.
Анохин предложил рассматривать функциональную организацию с точки зрения функциональных систем. Функциональная система – самоорганизующееся, саморегулирующееся образование, избирательно объединяющее различные элементы для достижения полезного приспособительного результата.
Выделяются два типа функциональных систем:
1. Системы первого типа обеспечивают гомеостаз за счёт внутренних (уже имеющихся) ресурсов организма, не выходя за его пределы (напр. кровяное давление)
2 .Системы второго типа поддерживают гомеостаз за счёт изменения поведения, взаимодействия с внешним миром, и лежат в основе различных типов поведения
Системообразующим фактором является результат (до рождения плод получает кислород из материнской крови (легочный круг не функционирует, спавшиеся легкие). Во время рождения нейроны дыхательного центра вызывают сокращение диафрагмы – легкие наполняются воздухом). Российский физиолог, основоположник теории функциональных систем Петр Кузьмич Анохин (1898-1974) выдвинул идею: решающий и единственный фактор — результат функционирования системы, который, будучи недостаточным, активно влияет на отбор именно тех степеней свободы из компонентов системы, которые при их интегрировании определяют дальнейшее получение полноценного результата. Формирование системы подчинено получению определенного полезного результата, а недостаточный результат может целиком реорганизовать систему и сформировать новую, с более совершенным взаимодействием компонентов, дающим достаточный результат. Что может быть более убедительным для доказательства справедливости положения о том, что результат является в самом деле центральным фактором системы. И вместе с тем что может быть очевиднее того, что не может быть понятия системы без ее полезного результата.
Принципы теории функциональных систем:
1. Принцип саморегуляции – при необходимости в организме автономно образуются функциональные системы, нормализующие параметры внутренней среды.
2. Принцип изоморфизма – равная организация функциональной системы.
3. Принцип мультипараметричности –
4. Принцип иерархичности – взаимодействующие элементы - кто-то доминирует, кто-то подчиняется (дыхание ровное, когда разговариваем, чихаем – неровное).
5. Принцип взаимосодействия – взаимосодействие направлено на положительный результат взаимодействия функциональных систем
6. Принцип системогенеза – функциональные системы созревают, они не готовы при рождении, причем созревают гетерохромно (неодновременно). Фактор, вызывающий созревание – результат деятельности самой системы.
2. Ионные механизмы потенциала покоя. Калиевый равновесный потенциал, формула Нернста.
МП, или потенциал покоя – это разность потенциалов между наружной и внутренней поверхностями мембраны клетки в условиях покоя. Внутренняя сторона мембраны клетки заряжена отрицательно по отношению к наружной, т.е. клеточная мембрана поляризована. В среднем у клеток возбудимых тканей он достигает 50-80мВ, со знаком "-" внутри клетки. Обусловлен преимущественно ионами калия. В клетках возбудимых тканей концентрация ионов калия достигает 150ммоль/л, в среде – 4-5ммоль (ионов калия намного больше в клетке, чем в среде). Поэтому по градиенту концентрации калий может выходить из клетки, и это происходит с участием калиевых каналов, часть которых открыта в условиях покоя. В результате из-за того, что мембрана непроницаема для анионов клетки (глутамат, аспарат, органические фосфаты), на внутренней поверхности клетки образуется избыток отрицательно заряженных частиц, а на наружной – избыток положительно заряженных частиц. Возникает разность потенциалов. Эта ситуация описывается уравнением Нернста.
Происхождение потенциала покоя.
Внутри
клетки концентрация ионов калия больше,
чем снаружи. Ионы калия компенсируют
заряд отрицательных ионов хлора и
других ионов органических молекул,
которые содержатся внутри клетки. С
другой стороны, из клетки постоянно с
помощью активного транспорта выводятся
ионы натрия. Поток ионов калия направлен
через калиевый канал из клетки.
Непроникающие анионы остаются в
цитоплазме, обеспечивая появление
разности потенциалов через мембрану,
и поэтому внутренняя сторона мембраны
заряжается отрицательно, что препятствует
дальнейшему выходу этих ионов. Эта
разность потенциалов препятствует
выходу ионов калия из клетки, и при
некотором ее значении наступает
равновесие ионных потоков. Возникает
равновесный
потенциал
, величина которого рассчитывается по
формуле Нернста:
 ,
где Ек
– равновесный потенциал для ионов
калия, R
– газовая постоянная, Т – абсолютная
температура, F
– число Фарадея (произведению Авогадро
числа NA
на элементарный электрический заряд
е (заряд электрона), n
– валентность К+(1),
,
где Ек
– равновесный потенциал для ионов
калия, R
– газовая постоянная, Т – абсолютная
температура, F
– число Фарадея (произведению Авогадро
числа NA
на элементарный электрический заряд
е (заряд электрона), n
– валентность К+(1),
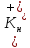 -
- – наружная и внутренняя концентрация
ионов калия. Если перейти от натуральных
логарифмов к десятичным и подставить
в уравнение числовые значения констант,
то уравнение примет вид:
– наружная и внутренняя концентрация
ионов калия. Если перейти от натуральных
логарифмов к десятичным и подставить
в уравнение числовые значения констант,
то уравнение примет вид:
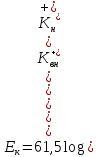 ,
при 37°С.
,
при 37°С.
Чем выше концентрация калия в среде – тем меньше это отношение, тем меньше величина мембранного потенциала. Однако расчетная величина, как правило, ниже реальной. Например, по расчетам МП должен быть -90мВ, а реально -70мВ. Это расхождение обусловлено тем, что ионы натрия и хлора тоже вносят свой вклад в создание МП. В частности, известно, что натрия больше в среде (140ммоль/л против 14ммоль/л внутриклеточной). Поэтому натрий может войти в клетку. Но большая часть натриевых каналов в условиях покоя закрыта. Поэтому в клетку входит лишь небольшая часть ионов натрия. Но и этого достаточно, чтобы хотя бы частично компенсировать избыток анионов. Ионы хлора, наоборот, входят в клетку (частично) и вносят отрицательные заряды. В итоге величина мембранного потенциала определяется в основном калием, а также натрием и хлором. Формула Гольдмана-Ходжкина описывает это явление.
Для того, чтобы МП поддерживался на постоянном уровне, необходимо поддержание ионного гетерогенитета – ионной асимметрии. Для этого, в частности, служит калий-натриевый насос (и хлорный), который восстанавливает ионную асимметрию, особенно после акта возбуждения. Доказательством калиевой природы МП является наличие зависимости: чем выше концентрация калия в среде, тем меньше величина МП. Для дальнейшего изложения важно понятие: деполяризация (уменьшение МП, например, от -90 мВ до -70 мВ) и гиперполяризация – противоположное явление.
Величина покоя будет определять уровень возбудимости. Возбудимость – способность отвечать на действие раздражителей изменением мембранного потенциала (обладают нейроны, мышечная ткань, секреторный эпителий).
Физиологические колебания потенциала покоя.
Сдвиги мембранного потенциала, вызываемые электрическим током, называются электротоническими потенциалами. При действии электрического тока подпороговой силы повышается (пропорционально силе тока) натриевая проницаемость мембраны, возникает явление деполяризации. Колебания мембранного потенциала вблизи критического уровня деполяризации называется локальным ответом. Локальный ответ не распространяется по мембране, суммируется при повторном действии раздражения, ему характерна повышенная возбудимость мембраны. При действии раздражителей на мембрану может возникнуть противоположное явление, т.е. ее гиперполяризация. При гиперполяризации повышается проницаемость мембраны для ионов калия и снижается ее возбудимость. Локальные измерения мембранного потенциала в виде деполяризации и гиперполяризации в физиологических
3. Локальный потенциал, изменение ионной проводимости мембраны при де- и реполяризации. Потенциал действия, мера возбудимости, порог, критический уровень деполяризации, овершут, гиперполяризация. Изменение возбудимости мембраны в процессе развития потенциала действия.
Локальный потенциал (ЛП) - это местное нераспространяющееся подпороговое возбуждение, существующее в пределах от потенциала покоя (-70 мВ) до критического уровня деполяризации (-50 мВ).
В случае превышения критического уровня деполяризации локальный потенциал переходит в потенциал действия и порождает нервный импульс.
Критический уровень деполяризации (КУД) - это такой уровень электрического потенциала мембраны возбудимой клетки, от которого локальный потенциал переходит в потенциал действия. В основе этого явления лежит самонарастающее открытие потенциал-управляемых ионных каналов для натрия под действием нарастающей деполяризации. КУД обычно составляет -50 мВ
Деполяризация обусловлена увеличением проницаемости мембраны для ионов натрия. При раздражении мембрана деполяризуется, что открывает натриевые каналы и создает поток ионов натрия внутрь клетки. Это дополнительно усиливает деполяризацию, увеличивает число открытых натриевых каналов и повышает натриевую проницаемость. Реполяризация закрывает натриевые каналы, что снижает натриевую проницаемость мембраны.
Потенциал действия – это кратковременное изменение разности потенциала между наружно и внутренней поверхностями мембраны (или между двумя точками ткани), возникающее в момент возбуждения. При регистрации потенциалы действия с помощью микроэлектродной техники наблюдается типичный пикообразный потенциал. В нем выделяют следующие фазы или компаненты:
1. Локальный ответ – начальный этап деполяризации.
2. Фазу деполяризации – быстрое снижение мембранного потенциала до нуля и перезарядка мембраны (реверсия, или овершут).
3. Фазу реполяризации – восстановление исходного уровня мембранного потенциала; в ней выделяют фазу быстрой реполяризации и фазу медленной реполяризации; в свою очередь, фаза медленной реполяризации представлена следовыми процессами (потенциалами):следовая негативность (следовая деполяризация) и следовая позитивность (следовая гиперполяризация). Амплитудно-временные характеристики потенциала действия нерва, скелетной мышцы таковы: амплитуда потенциала действия 140-150мВ; длительность пика потенциала действия (фаза деполяризации + фаза реполяризации) составляет 1-2 мс, длительных следовых потенциалов – 10-50 мс.
При исследовании ПД аксонов и сомы нервной клетки, ПД скелетной мышцы было установлено, что фаза деполяризации обусловлена значительным повышением проницаемости для ионов натрия, которые входят в клетку в начале процесса возбуждения и таким образом уменьшают существующую разность потенциалов (деполяризация). Процесс этот имеет регенеративную природу – чем выше степень деполяризации, тем выше становится проницаемость натриевых каналов, тем больше входит ионов натрия в клетку и тем выше степень деполяризации. В этот период происходит не только снижение разности потенциалов до нуля, но и изменение поляризованности мембраны – на высоте пика ПД внутренняя поверхность мембраны заряжена положительно по отношению к наружной (явление реверсии, или овершута). Однако бесконечно этот процесс идти не может: в результате закрытия инактивационных ворот натриевые каналы закрываются, и приток натрия в клетку прекращается. Затем наступает фаза реполяризации. Она связана с увеличением выхода из клетки ионов калия. Это происходит за счет того, что в результате деполяризации большая часть калиевых каналов, которые в условиях покоя были закрыты, открываются и "+" заряды уходят за пределы клетки. Вначале этот процесс идет очень быстро, потом – медленно, поэтому фаза реполяризации вначале протекает быстро (нисходящая часть пика ПД), а потом медленно (следовая негативность). Этот же процесс лежит в основе фазы следовой гиперполяризации.
Если на клетку действуют раздражители – процесс возбуждения. Процесс уменьшения электроотрицательности - деполяризация – при действии раздражителя. Восстановление мембранного потенциала после деполяризации – реполяризация. С увеличением раздражителя деполяризация развивается быстрее и на большую величину. Потенциал действия означает возбуждение. Деполяризация обусловлена входом ионов натрия внутрь клетки по градиенты концентрации через натриевые каналы. Критический уровень деполяризации — величина мембранного потенциала, при достижении которой возникает потенциал действия. Реполяризация обусловлена выходом ионов калия из клетки по градиенту концентрации через калиевые каналы. Начинается потому, что открываются калиевые каналы. Гиперполяризация возникает из-за того, что не успели вовремя закрыться калиевые каналы. При гиперполяризации повышается проницаемость мембраны для ионов калия и снижается ее возбудимость. Мерой возбудимости является минимальная сила раздражителя, вызывающего возбуждение. Такой раздражитель называется пороговым. Между порогом и возбудимостью существует обратная связь (зависимость). Чем больше порог, тем меньше возбудимость и наоборот. Возбудимость – непостоянная величина. 3 уровня возбудимости: нормальная, повышенная (=экзальтация), рефрактерность (=невозбудимость или сниженная). Абсолютная рефрактерность определяет свойство лабильности – часто возбуждаться, быстро реагировать на раздражителя и обрабатывать информацию. Для поддержания разницы концентрации ионов натрия и калия по обе строны мембраны существует транспортная система натрий-калиевая АТФаза.
4. Свойства порогового раздражителя: закон "все или ничего", зависимость между силой и длительностью порогового раздражителя (хронаксия), аккомодация. Характеристика ионных каналов Na+, K+ и Na+/K+-АТФазы.
Закон "все или ничего": под пороговые раздражители не вызывают ответной реакции ("ничего"), на пороговые раздражители возникает максимальная ответная реакция ("все"). По закону "все или ничего" сокращаются сердечная мышца и одиночное мышечное волокно. Закон "все или ничего" не абсолютен. Во-первых, на раздражители подпороговой силы не возникает видимой ответной реакции, но в ткани происходят изменения мембранного потенциала покоя в виде возникновения местного возбуждения (локального ответа). Во-вторых, сердечная мышца, растянутая кровью, при наполнении ею камер сердца, реагирует по закону "все или ничего", но амплитуда ее сокращения будет больше по сравнению с сокращением сердечной мышцы, не растянутой кровью.
Закон силы-длительности: раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать для возникновения возбуждения.
Исследования зависимости силы-длительности показали, что последняя имеет гиперболический характер. Из этого следует, что ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Причиной такой' зависимости является мембранная емкость. Очень "короткие" токи просто не успевают разрядить эту емкость до критического уровня деполяризации. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого действует ток, равный реобазе, и вызывает возбуждение, называется полезным временем.
Закон раздражения Дюбуа-Реймона (аккомодации): раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени. При действии медленно нарастающего раздражителя возбуждение не возникает, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации. Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает. Способность к аккомодации различных структур неодинакова. Наиболее .высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
Закон силы. Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным – пороговым или выше порогового.
Закон времени. Раздражитель, вызывающий возбуждение, должен быть достаточно длительным, воздействовать на ткань некоторое время, чтобы вызвать возбуждение.
Закон градиента. Для того, чтобы раздражитель вызвал возбуждение, он должен нарастать достаточно быстро. Если раздражитель нарастает медленно, то в силу развития аккомодации (инактивации натриевых каналов), происходит повышение порога раздражения, поэтому для получения возбуждения величина стимула должна быть больше, чем если бы он нарастал мгновенно. Гиперболический характер (является обратно-пропорциональная зависимость).
Характеристика ионных каналов.
|
Свойство |
Na+ |
K+ |
Na+/K+-АТФаза |
|
1. Избирательность транспорта |
Na+ |
K+ |
3Na+=2K+ |
|
2. Направление |
по градиенту концентрации |
против градиента концентрации | |
|
3. потенциалзависимость |
+ |
+ |
- |
|
4. энергозависимоть |
- |
- |
+ АТФ |
|
5. ингибиторы (блокаторы) транспорта |
Тетродотоксин (рыба фуга) |
Тетраэтиламмоний |
строфантин |