

Polnyj_kurs_lekcij
.pdfbiokhimija.ru |
Тимин О.А. Лекции по биологической химии |
201 |
|
|
|
Однако надо иметь в виду и пятую, глицеральдегидфосфат-дегидрогеназную, реакцию, из которой выходит НАДН. Если условия анаэробные, то он используется в лактатдегидрогеназной реакции, где окисляется для образования лактата и в получении АТФ не участвует.
Аэробное окисление
Если в клетке имеется кислород, то НАДН из шестой реакции гликолиза направляется в митохондрию, на процессы окислительного фосфорилирования. Там его окисление приносит дивиденды в виде трех молекул АТФ.

Строение и обмен углеводов |
202 |
|
|
Образовавшийся в гликолизе пируват в аэробных условиях превращается в ПВК-дегидрогеназном комплексе в ацетил-S-КоА, при этом образуется 1 молекула НАДН.
Ацетил-S-КоА вовлекается в ЦТК и, окисляясь, дает 3 молекулы НАДН, 1 молекулу ФАДН2, 1 молекулу ГТФ. Молекулы НАДН и ФАДН2 движутся в дыхательную цепь, где при их окислении в сумме образуется 11 молекул АТФ. В целом при сгорании одной ацетогруппы в ЦТК образуется 12 молекул АТФ.
Суммируя результаты окисления "гликолитического" и "пируватдегидрогеназного" НАДН, "гликолитический" АТФ, энергетический выход ЦТК и умножая все на 2, получаем
38 молекул АТФ.
Общее уравнение аэробного окисления глюкозы имеет вид:
C6H12O6 + 6 O2 + 38 АДФ + 38 Фн → 6 CO2 + 44 H2О + 38 АТФ
ЭФФЕКТ ПАСТЕРА
Эффект Пастера – это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода.
Биохимический механизм эффекта Пастера заключается в конкуренции между пиру-
ватдегидрогеназой, превращающей пируват в ацетил-S-КоА, и лактатдегидрогеназой, пре-
вращающей пируват в лактат.
При отсутствии кислорода внутримитохондриальные процессы дыхания не идут, моментально накапливающийся НАДН тормозит цикл трикарбоновых кислот и накапливающийся ацетил-S-КоА дополнительно ингибирует ПВК-дегидрогеназу. В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную.
При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.

biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
203 |
|
|
|
Иллюстрацией к сказанному служит отличие изоферментов лактатдегидрогеназ (ЛДГ) друг от друга. Сердечный изофермент ЛДГ-1 обладает высоким сродством к молочной кислоте и "стремится" поднять концентрацию пирувата с целью его включения в ЦТК и получения энергии для деятельности миокарда. Большое количество митохондрий и поступление сюда лактата из других органов обеспечивает работу сердца при аэробных условиях. При нехватке кислорода свойства ЛДГ-1 не изменятся, он по-прежнему будет сдвигать реакцию в
сторону продукции пировиноградной кислоты, и накопление пирувата вызовет необратимые повреждения кардиомиоцита.
В тоже время изофермент скелетной мышцы ЛДГ-5 обладает высоким сродством к пирувату, при отсутствии кислорода в клетке быстро и эффективно превращает его в лактат, легко проникающий сквозь мембраны и удаляемый из мышцы.
Таким образом, в анаэробных условиях сильнее будет страдать сердечная мышца, что, собственно говоря, и наблюдается в медицинской практике.
Важно то, что пировиноградная кислота является для клетки токсичным веществом, и клетке необходимо избавиться от нее каким угодно образом. Так как через мембраны она не проходит, то обезвреживание достигается превращением пирувата 1) в лактат; 2) в ацетил-S-КоА; 3) в аланин (см "Аланинаминотрансфераза"), 4) в оксалоацетат.
ЧЕЛНОЧНЫЕ СИСТЕМЫ
Молекулы НАДН, образованные в шестой реакции гликолиза, в зависимости от наличия кислорода имеют, как минимум, два пути своего дальнейшего превращения:
oлибо остаться в цитозоле и вступить в лактатдегидрогеназную реакцию (анаэробные условия),
o либо проникнуть в митохондрию и окислиться в дыхательной цепи (аэробные условия), Так как сама молекула НАДН через митохондриальную мембрану не проходит, то су-
ществуют системы, принимающие этот водород в цитоплазме и отдающие его в матриксе митохондрий. Они называются челночные системы.
Определены две основные челночные системы – глицеролфосфатная и малат-
аспартатная.
Глицеролфосфатный челночный механизм
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глице- рол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол- 3-фосфат, поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии.
Работа глицеролфосфатного челночного механизма актуальна при необходимости получить энергию из глюкозы при работе клетки.
Однако, если в клетке имеется избыток энергии (состояние покоя, после еды) то гли- церол-3 фосфат будет использоваться в цитозоле для синтеза жиров (см "Синтез триацилглицеролов").

Строение и обмен углеводов |
204 |
|
|
Малат-аспартатный челночный механизм
Этот механизм более сложен: постоянно идущие в цитоплазме реакции трансаминирования аспартата поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и "гликолитического" НАДН восстанавливается до яблочной кислоты. Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он аминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.

biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
205 |
|
|
|
|
|
|
ГЛЮКОНЕОГЕНЕЗ
В клетках организма всегда существует потребность в глюкозе:
o для эритроцитов глюкоза является единственным источником энергии,
oнервная ткань потребляет около 120 г глюкозы в сутки, что составляет до 70% глюкозы образующейся в печени, притом эта величина практически не зависит от интенсивности ее работы. Только в экстремальных ситуациях (длительное голодание) она
способна получать энергию из неуглеводных источников,
oглюкоза играет весомую роль для поддержания необходимых концентраций метаболитов цикла трикарбоновых кислот (в первую очередь оксалоацетата).
Таким образом, при определенных ситуациях – при низком содержании углеводов в пище, голодании, длительной физической работе – организм должен иметь возможность получить глюкозу. Это достигается процессом глюконеогенеза.
Глюконеогенез – это синтез глюкозы из неуглеводных компонентов: лактата, пирувата, глицерола, кетокислот цикла Кребса и других кетокислот, из аминокислот. Процесс вклю-
чает все обратимые реакции гликолиза, и особые обходные пути, т.е. он не полностью по-
вторяет реакции окисления глюкозы. Его реакции способны идти во всех тканях, кроме последней глюкозо-6-фосфатазной реакции, которая идет только в печени и почках. Поэтому, строго говоря, глюконеогенез идет только в этих двух органах.
Как указывалось ранее, в гликолизе существуют три необратимые реакции: пируват-
киназная (десятая), фосфофруктокиназная (третья) и гексокиназная (первая). На этих ре-
акциях существуют энергетические барьеры, которые обходятся с помощью дополнительных реакций.
Все аминокислоты, кроме кетогенных лейцина и лизина, способны участвовать в синтезе глюкозы. Углеродные атомы некоторых из них – глюкогенных – полностью включаются в молекулу глюкозы, некоторых – смешанных – частично.
Кроме получения глюкозы, глюконеогенез обеспечивает и уборку "шлаков" – лактата, постоянно образуемого в эритроцитах или при мышечной работе и глицерола, являющегося продуктом липолиза в жировой ткани.
ОБХОД ДЕСЯТОЙ РЕАКЦИИ ГЛИКОЛИЗА
На этом этапе глюконеогенеза работают два основных фермента – в митохондриях пи-
руваткарбоксилаза и в цитозоле фосфоенолпируват-карбоксикиназа.
Вмитохондриях пируваткарбоксилаза превращает пировиноградную кислоту в оксалоацетат. Необходимо отметить, что эта реакция идет в клетке постоянно, являясь анаплеротической (пополняюшей) реакцией ЦТК.
Далее оксалоацетат должен попасть в цитозоль и превратиться в фосфоенолпируват. Однако дело осложняется непроницаемостью мембраны для оксалоацетата. Зато через мембрану может пройти малат, предшественник оксалоацетата по ЦТК. Повернуть малатдегид-
рогеназную реакцию ЦТК вспять позволяет избыток НАДН в митохондриях. НАДН поступает из β-окисления жирных кислот, активируемого в условиях недостаточности глюкозы в гепатоците.
Врезультате малат накапливается, выходит в цитозоль и здесь превращается обратно в оксалоацетат.
Вцитоплазме фосфоенолпируват-карбоксикиназа осуществляет превращение оксалоацетата в фосфоенолпируват, для реакции требуется энергия ГТФ. От молекулы отщепляется тот же углерод, что и присоединяется.

Строение и обмен углеводов |
206 |
|
|
ОБХОД ТРЕТЬЕЙ РЕАКЦИИ ГЛИКОЛИЗА
Второе препятствие на пути синтеза глюкозы – фосфофруктокиназная реакция – преодолевается с помощью фермента фруктозо-1,6-дифосфатазы. Этот фермент есть в почках, печени, поперечно-полосатых мышцах. Таким образом, эти ткани способны синтезировать фруктозо-6-фосфат и глюкозо-6-фосфат.
ОБХОД ПЕРВОЙ РЕАКЦИИ ГЛИКОЛИЗА
Последняя реакция катализируется глюкозо-6-фосфатазой. Она имеется только в печени и почках, следовательно, только эти ткани могут продуцировать свободную глюкозу.
ГЛЮКОЗО- ЛАКТАТНЫЙ И ГЛЮКОЗО- АЛАНИНОВЫЙ ЦИКЛЫ
Глюкозо-лактатный цикл (цикл Кори) – это связь глюконеогенеза в печени и образования лактата в эритроцитах или мышцах из глюкозы. В эритроцитах молочная кислота об-

biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
207 |
|
|
|
разуется непрерывно, так как для них анаэробный гликолиз является единственным способом образования энергии.
В скелетных мышцах высокое накопление молочной кислоты является следствием гликолиза при очень интенсивной, субмаксимальной мощности, работе. Но даже при работе низкой и средней интенсивности в скелетной мышце образуется некоторое количество лактата. Утилизоваться он может только одним способом – превратиться в пировиноградную кислоту. Однако, сама мышечная клетка ни при работе, ни во время отдыха не способна превратить лактат в пируват из-за особенностей изофермента ЛДГ-5.
Поэтому во время и после нагрузки (при восстановлении) лактат удаляется из мышцы. Это происходит довольно быстро, всего через 0,5-1,5 часа в мышце лактата уже нет. Малая часть молочной кислоты выводится с мочой. Большая часть лактата захватывается гепатоцитами, окисляется в пировиноградную кислоту и вступает на путь глюконеогенеза. Полученная глюкоза используется самим гепатоцитом или выходит в кровь, пополняя запасы в мышце или других клетках.
Целью глюкозо-аланинового цикла также является уборка пирувата, но, кроме этого решается еще одна немаловажная задача – уборка лишнего азота из мышцы. При мышечной работе и в покое в миоците распадаются белки и образуемые аминокислоты трансаминируются с α-кетоглутаратом. Полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой азота и пирувата из мышцы в печень. В гепатоците идет обратная реакция трансаминирования, аминогруппа передается на синтез мочевины, пируват используется для синтеза глюкозы
Глюкоза, образованная в печени из лактата или аланина, возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена. Также она может распределиться по другим органам.
Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время голодания, когда мышечные белки распадаются и многие аминокислоты используются в качестве источника энергии, а их азот необходимо доставить в печень.
Строение и обмен углеводов |
208 |
|
|
ВЗАИМОСВЯЗЬ ГЛИКОЛИЗА И ГЛЮКОНЕОГЕНЕЗА
Так как синтез и окисление глюкозы являются крайне важными для существования клетки (гликолиз) и всего организма (глюконеогенез), то регуляция этих процессов отвечает запросам органов и тканей при различных условиях существования.
Поскольку гликолитическое окисление глюкозы:
oявляется способом получения энергии и в аэробных и в анаэробных условиях, то оно непрерывно происходит во всех клетках и, конечно, должно и будет активировано
при усилении работы клетки, например, сокращение миоцита, движение нейтрофилов;
o является источником глицерола и ацетил-SКоА, используемых для синтеза жиров в гепатоцитах и адипоцитах, то такое окисление активируется при избытке глюкозы в клетке.
Глюконеогенез, как образование глюкозы в печени из неуглеводных компонентов, необходим:
oпри гипогликемии во время мышечной нагрузки – синтез из молочной кислоты, поступающей из мышц, из глицерола, образующегося при мобилизации жиров;
oпри гипогликемии при голодании – синтез из аминокислот, образующихся при катаболизме белков.
Таким образом, при голодании или физической нагрузке глюконеогенез, идущий в печени, обеспечивает глюкозой все остальные органы (эритроциты, нервная ткань, мышцы и др.), в которых активен гликолиз и другие процессы, производящие энергию. Наличие глюкозы в указанных клетках необходимо, чтобы поддержать концентрацию оксалоацетата и обеспечить сгорание ацетил-SКоА в цикле трикарбоновых кислот.
РЕГУЛЯЦИЯ ГЛИКОЛИЗА И ГЛЮКОНЕОГЕНЕЗА
Вцелом можно выделить два способа регуляции гликолиза и глюконеогенеза: гормональная и метаболическая, т.е. при помощи промежуточных или конечных продуктов обмена глюкозы.
Существуют три основных участка, на которых происходит регуляция: o первая реакция гликолиза,
o третья реакция гликолиза и обратимая ей, o десятая реакция гликолиза и обратимые ей.
Регуляция глюконеогенеза
Гормональная активация глюконеогенеза осуществляется глюкокортикоидами, кото-
рые увеличивают синтез пируват-карбоксилазы, фосфоенолпируваткарбоксикиназы,
фруктозо-1,6-дифосфатазы. Глюкагон стимулирует те же самые ферменты через аденилатциклазный механизм путем фосфорилирования.
Кроме гормонов, имеются метаболическая регуляция, при которой аллостерически активируется пируваткарбоксилаза при помощи ацетил-S-КоА, фруктозо-1,6-дифосфатаза
при участии АТФ.
Регуляция гликолиза
Гликолиз стимулируется инсулином, повышающим количество молекул гексокиназы,
фосфофруктокиназы, пируваткиназы.
В печени активность глюкокиназы регулируется гормонами: активацию вызывает инсулин и андрогены, подавляют ее активность глюкокортикоиды и эстрогены.
К метаболической регуляции наиболее чувствительна фосфофруктокиназа. Она активируется АМФ и собственным субстратом, ингибируется – АТФ, лимонной кислотой, жирными кислотами. Пируваткиназа активируется фруктозо-1,6-дифосфатом. Гексокиназа

biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
209 |
|
|
|
клеток (кроме гепатоцитов) ингибируется продуктом собственной реакции – глюкозо-6- фосфатом.
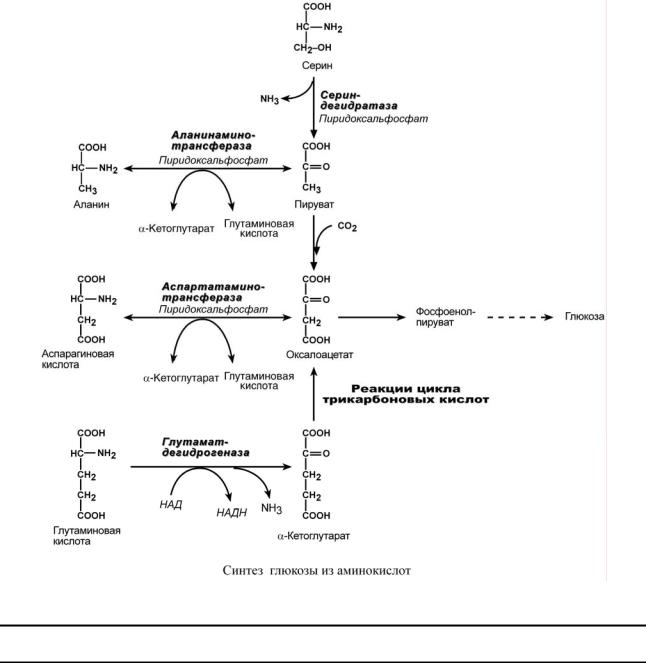
СИНТЕЗ ГЛЮКОЗЫ ИЗ АМИНОКИСЛОТ
Ряд аминокислот являются глюкогенными, то есть их углеродные скелеты в той или иной степени способны включаться в состав глюкозы. Такими является большинство аминокислот, кроме лейцина и лизина, углероды которых никогда не участвуют в синтезе углеводов.
В качестве примера синтеза глюкозы из аминокислот рассмотрим участие в этом процессе глутамата, аспартата, серина и аланина.
Аспарагиновая кислота (после реакции трансаминирования) и глутаминовая кисло-
та (после дезаминирования) превращаются в метаболиты ЦТК, соответственно, в оксалоацетат и α-кетоглутарат.
Аланин, трансаминируясь, образует пировиноградную кислоту, которая способна карбоксилироваться до оксалоацетата. Оксалоацетат, являясь первым элементом в процессе глюконеогенеза, далее включается в состав глюкозы.
Серин в трехступенчатой реакции под воздействием сериндегидратазы теряет аминогруппу и превращается в пируват, который вступает в глюконеогенез.
Строение и обмен углеводов |
210 |
|
|
МЕТАБОЛИЗМ ЭТАНОЛА
В организм поступают и в результате метаболизма образуются в клетках головного мозга и печени, в других тканях, при жизнедеятельности микрофлоры кишечника различные спирты (алифатической, ароматической, стероидной природы, ретиноиды, фарнезол и др.) и альдегиды (ароматические, алифатические, продукты пероксидации липидов и др.), которые могут являться промежуточными метаболитами или конечными продуктами. Взаимопревращение спиртов и альдегидов осуществляют алкогольдегидрогеназы. Их существует 6 классов, в каждом классе имеются многочисленные изоферменты, обнаруженные во многих тканях.
С медицинской и социальной точки зрения определенный интерес вызывает метаболизм этилового спирта в организме человека.