

Синтез атф регулируется
Работа дыхательных ферментов регулируется с помощью эффекта, который получил название дыхательный контроль.
Дыхательный контроль
Дыхательный контроль – это влияние электрохимического градиента на объём движения электронов по дыхательной цепи. В свою очередь, величина градиента напрямую зависит от соотношения АТФ и АДФ, количественная сумма которых в клетке практически постоянна ([АТФ] + [АДФ] = const). Реакции катаболизма направлены на поддержание постоянно высокого уровня АТФ и низкого АДФ.
Возрастание протонного градиента возникает при снижении количества АДФ и накоплении АТФ (состояние покоя), т.е. ионы Н+ не проникают в матрикс митохондрии. При этом ингибирующее влияние градиента усиливается и продвижение электронов по цепи уменьшается. Ферментные комплексы остаются в восстановленном состоянии. Следствием является уменьшение окисления НАДН и ФАДН2 на I и II комплексах и как следствие замедление катаболизма в клетке.
Снижение протонного градиента возникает при снижении резервов АТФ и избытке АДФ. В этом случае активно работает АТФ-синтаза и через канал Fо проходят в матрикс ионы Н+. При этом градиент, естественно, снижается, поток электронов возрастает, в результате повышается выкачивание ионов Н+ в межмембранное пространство и синтез АТФ. Ферментные комплексы I и II усиливают окисление НАДН и ФАДН2 (как источников электронов) и снимается ингибирующее влияние НАДН на цикл лимонной кислоты и пируватдегидрогеназный комплекс. Как итог – активируются реакции катаболизма углеводов и жиров.
На клеточное дыхание также могут влиять: Гипоэнергетические состояния
Причиной гипоэнергетических состояний может быть следующее:
гиповитаминозы экзогенные и/или эндогенные – снижается скорость и эффективность окислительных реакций. Возникает обычно при нехватке витаминов – В1, В2, В3, В5, В6, В7 и аскорбиновой кислоты;
дефицит белка в пище – снижается синтез всех ферментов и ферментов катаболизма в частности,
снижение потребления углеводов и липидов как основных источников энергии,
дефицит кислорода – отсутствие акцептора для электронов вызывает "переполнение" дыхательных ферментов, повышение электрохимического градиента, накопление НАДН и ФАДН2 в клетке и прекращение катаболизма,
дефицит железа – компонента цитохромов, миоглобина и гемоглобина, меди – компонента цитохромоксидазы, магния – стабилизатора молекулы АТФ.
Разобщители окисления и фосфорилирования
При определённых условиях протоны могут возвращаться в матрикс без синтеза АТФ. Этот процесс известен как утечка электронов или митохондриальное разобщение и связано с диффузией протонов в матрикс.
Процесс осуществляется с помощью каналов, образованных белком термогенином или (thermogenin или UCP1), составляющий у новорожденных до 15% от всех белков митохондрий. Термогенин небольшой белок (33 kDa) открыт в 1973 году. Вначале был найден в бурой жировой ткани и был ответственен за термогенез без дрожания. Наибольшее количество бурой жировой ткани имеют новорожденные, его количество значительно падает с возрастом.
При охлаждении организма бурые адипоциты получают сигналы по симпатическим нервам, и в них активируется расщепление жира – липолиз. Окисление жиров приводит к получению большого количества НАДН и ФАДН2, активизации работы дыхательной цепи и возрастанию электрохимического градиента. Благодаря термогенину большая часть энергии ионов водорода рассеивается в виде тепла, подогревая протекающую через ткань кровь и обеспечивая поддержание температуры тела при охлаждении.
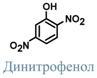
Вещества, которые снижают величину электрохимического градиента, приводя к увеличению объёма электронов движущихся по ферментам дыхательной цепи, уменьшению синтеза АТФ и возрастанию катаболизма - называются разобщители. Они разобщают (разъединяют) процессы окисления и фосфорилирования внутренней мембраны митохондрий. К разобщителям в первую очередь относят "протонофоры" – вещества переносящие ионы водорода. При этом одновременно уменьшаются оба компонента электрохимического градиента – электрический и химический, и энергия градиента рассеивается в виде тепла. Следствием эффекта протонофоров является возрастание катаболизма жиров и углеводов в клетке и во всем организме.
Классическим протонофором является динитрофенол, жирорастворимое соединение, присоединяющие ионы водорода на внешней поверхности внутренней митохондриальной мембраны и отдающие их на внутренней поверхности. Белок термогенин является физиологическим протонофором.
Кроме динитрофенола и термогенина протонофорами, к примеру, являются салицилаты, жирные кислоты и трийодтиронин.