

мов, приводящих к гипоксическому состоянию клетки тех органов, которые чаще всего используются для трансплантации (сердце, печень, почки), поскольку эти органы неизбежно проходят стадию гипоксии со времени их удаления из организма донора до момента имплантации в организм реципи-
ента (Kehrer J.P., Concepts related to the study of reactive oxygen and cardiac reperfusion injury, Free Rad. Res. Commun., 1986, 5, 305-317 и Granger D.N., Role xanthine oxidase and granulocytes in ischemia – reperfusion injury, Am. J. Physiol., 1988, 255, H1269).
Утилизация молочной кислоты
Несмотря на отрицательные эффекты избытка молочной кислоты, образующейся в ходе гликолиза на фоне гипоксических проявлений, это соединение достаточно эффективно утилизируется печенью с последующим синтезом глюкозы.
Действительно, основным исходным соединением, которое используется в глюконеогенезе является молочная кислота, которая образуется при активной работе скелетных мышц. В сокращающейся мышце при анаэробных условиях скорость образования пирувата в ходе гликолитического расщепления глюкозы превышает скорость его поступления в митохондрии, и, следовательно, скорость последующего окисления этого интермедиата в цикле трикарбоновых кислот. Кроме того, при интенсивно протекающем гликолизе скорость образования NADH под действием глицеральдегид-3-фосфат- дегидрогеназы выше, чем скорость его окисления в дыхательной цепи. Отсюда следует, что интенсивно протекающий гликолиз требует постоянного окисления NADH в NAD+. Генерирование NAD+ осуществляется лактатдегидрогеназой, которая восстанавливая пировиноградную кислоту в молочную, окисляет NADH в NAD+.
Образование молочной кислоты – это тупик в метаболизме. Дальнейшие метаболические превращения молочной кислоты возможны только в случае повторного образования пирувата из лактата:
Исходя из сказанного выше, очевидно, что единственной целью восстановления пирувата в молочную кислоту является регенерация окисленной формы никотинамид-аденинидинуклеотида – NAD+, необходимого для непрерывного протекания гликолитического пути расщепления глюкозы в ак-
51
тивной скелетной мышце. В свою очередь, процесс образования молочной кислоты в мышцах перекладывает часть метаболической нагрузки на печень.
Ранее обсуждалось, что плазматические мембраны большинства клеток обладают высокой проницаемостью для молочной и пировиноградной кислот. Оба соединения диффундируют из активной скелетной мышцы в кровь и переносятся в печень. При этом количество переносимого лактата существенно превышает количество пировиноградной кислоты из-за высокого значения отношения концентраций [NADH]/[NAD+] в сокращающихся мышцах. Молочная кислота, поступившая в печень, снова окисляется в гепатоцитах до пирувата вследствие низкого отношения концентраций [NADH]/[NAD+] в цитозоле этих клеток. Образовавшийся в печени пируват затем превращается в глюкозу по пути глюконеогенеза.
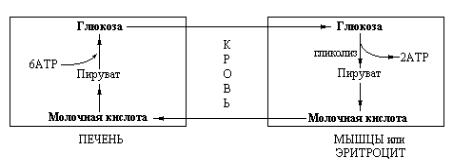
Таким образом, печень снабжает глюкозой сокращающиеся мышцы, которые, в свою очередь, генерируют АТР в результате гликолитического превращения глюкозы в молочную кислоту. Молочная кислота снова превращается в печени в глюкозу. Указанные взаимопревращения глюкозы и молочной кислоты составляют цикл Кори (рис. 4.2).
Рис. 4.2 Схема функционирования цикла Кори. Молочная кислота, образующаяся в активной скелетной мышце, превращается в печени в глюкозу. Этот цикл передает часть метаболического «груза» активной мышцы гепатоцитам.
Координированное функционирование описанных процессов облегчается различиями в каталитических свойствах лактатдегидрогеназ, локализованных в мышечной и печеночной тканях. Лактатдегидрогеназа представляет собой тетрамер, состоящий из субъединиц с молекулярной массой 35 000 дальтон, каждая. Однако субъединицы лактат-дегидрогеназы не идентичны: существует два вида полипептидных цепей, обозначаемых буквами М (muscle) и Н (hepatic), которые могут образовывать, что очевидно, пять форм фермента, называемых изоферментами. Наибольшим сродством к пирувату обладает изофермент М4, а изоформа Н4 характеризуется существенно более низким сродством к пировиноградной кислоте, остальные изоферменты обладают промежуточной активностью.
52