


ХОД РАБОТЫ
*Задание Определить концентрацию мочевины в сыворотке крови с использованием тест-набора клинического назначения
1. Готовят опытную, стандартную и холостую пробы в соответствии с таблицей:
|
|
|
|
|
|
|
Ингредиенты |
Опытная |
Стандартная |
Холостая |
|
|
|
проба, мл |
проба, мл |
проба, мл |
|
|
Сыворотка крови |
0.2 |
− |
− |
|
|
Мочевина, стандарт |
− |
0.2 |
− |
|
|
Дистиллир. вода |
0.8 |
0.8 |
0.5 |
|
|
ТХУ |
1.0 |
1.0 |
− |
|
|
|
|
|
|
|
|
|
|
|
|
|
2.Опытную и стандартную пробы перемешивают стеклянной палочкой, выдерживают 15 мин, затем центрифугируют при 1500 об/мин в течение 10 мин.
3.Из центрифужных пробирок опытной и стандартной проб отбирают по 0.5мл супернатанта, вносят по 5мл цветного реактива (к 0.5 мл холостой пробы также добавляют 5 мл цветного реагента) и содержимое пробирок перемешивают.
4.Пробирки помещают в кипящую водяную баню на 20 мин. Затем охлаждают 2-3 мин под струей воды.
5.Оптическую плотность опытной и стандартной проб измеряют при длине волны 540 нм, используя в качестве контроля холостую пробу.
6.Расчет концентрации мочевины в сыворотке крови производят по формуле:
Х= Еоп 16.65,
Ест
где Х - концентрация мочевины в сыворотке крови (мМ), Еоп – оптическая плотность опытной пробы, Ест – оптическая плотность стандартной пробы,
16.65 - концентрация мочевины в стандартном растворе (мМ).
Примечание: норма содержания мочевины в сыворотке крови составляет 2.5- 8.3 мМ.
Оформление работы
К занятию:
1. Кратко законспектировать теоретические материалы по лабораторной работе.
25
Во время занятия:
2.Описать этапы работы.
3.Построить калибровочные графики.
4.Оформить результаты измерений.
5.Сделать выводы.
26
Методические указания к лабораторному практикуму по курсу «Патобиохимия»
Тема:
Цель работы:
ЛАБОРАТОРНАЯ РАБОТА № 3
Определение перекисного гемолиза эритроцитов и уровня восстановленного глутатиона в эритроцитах крови человека
Определить уровень перекисного гемолиза эритроцитов. Определить соотношение концентраций восстановленной и окисленной форм глутатиона в эритроцитах крови человека
Оборудование и материалы:
•Центрифуга UNIVERSAL 30RF
•Пробирки центрифужные
•Спектрофотометр SOLAR
•Кюветы полистирольные
•Автоматические микропипетки
•Пробирки
•Штативы для пробирок
•Цилиндры мерные на 50-100мл
•Бумага фильтровальная
•Палочка стеклянная
Реактивы:
•Цельная кровь
•Натрий хлористый (NaCl), 17% раствор
•Калий-фосфатный буфер, pH7,4
•Вода дистиллированная
•Кровь цельная (соотношение с антикоагулянтом - 4:1)
•Осаждающий реактив:
метафосфорная кислота (HPO3), трилон Б, хлористый натрий (NaCl)
• Реактив Эллмана:
0.04% раствор 5,5,-дитиобис(2-нитробензойной кислоты) в 1% растворе цитрата натрия трехзамещенном
•Гидроортофосфат натрия (Na2HPO4), 0.3М раствор
•Глутатион восстановленный, стандартный раствор (5мМ)
•Физиологический раствор (NaCl, 0.9% раствор)
•Вода дистиллированная
27
ТЕОРЕТИЧЕСКАЯ ЧАСТЬ
Антиоксидантная система защиты клеток от действия продуктов перекисного окисления липидов
Процессы биологического окисления в клетках в норме представлены рядом метаболических (катаболических) путей, которые включают последовательные ферментативные реакции дегидрирования, то есть отщепления атомов водорода от окисляемого соединения, а кислород, как правило, присоединяется не к самому окисляемому веществу, а к высвобождаемым атомам водорода с образованием молекул воды. Вместе с тем, в разных клетках присутствуют также специфические ферменты или даже мультиферментные системы, которые, используя молекулярный кислород, способны непосредственно внедрять один (монооксигеназы) или два (диоксигеназы) атома кислорода в окисляемый субстрат.
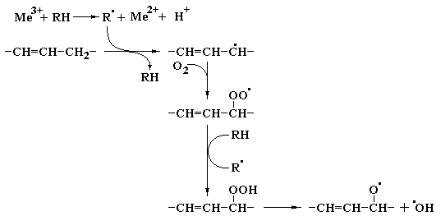
Наконец, под действием ряда ферментов или в ходе неферментативных реакций, катализируемых следовыми количествами переходных металлов, может происходить инициирование аутокаталитических цепных реакций, в которых основными субстратами выступают соединения, содержащие двойные связи, чередующиеся с метиленовыми мостиками. Такие особенности структуры характерны для природных полиненасыщенных жирных кислот. Приведенная ниже последовательность реакций включает регенерирование радикала R·, обеспечивающего развитие цепной реакции, и одновременное образование в присутствии кислорода органических гидроперекисей, которые могут распадаться на два кислородсодержащих радикала, вступающих в последующие реакции:
Указанную последовательность реакций можно также представить в виде следующих уравнений:
28
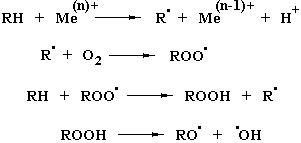
Перекисное окисление липидов (ПОЛ) в присутствии кислорода вызывает in vivo повреждение тканей, способствуя развитию опухолевых заболеваний. Процессы перекисного окисления in vivo могут катализироваться гемовыми белками и липоксигеназами, входящими в состав тромбоцитов, лейкоцитов, эритроцитов и других форменных элементов крови. Повреждающее действие инициируется свободными кислородсодержащими радикалами – ROO·, RO·, HO·, возникающими в ходе образования перекисей жирных кислот.
Одним из неблагоприятных последствий перекисного окисления липидов считается накопление малонового альдегида, образующегося в результате радикального разрыва полиеновых жирных кислот. Этот бифункциональный альдегид способен образовывать Шиффовы основания с аминогруппами белков, выступая в качестве внутри- и межмолекулярного сшивающего агента. В результате сшивки формируются нерастворимые липид-белковые ком-
плексы, называемые пигментами изнашивания.
Контроль за физиологическим уровнем продуктов ПОЛ осуществляет мощная естественная антиоксидантная система, включающая низкомолекулярные антиоксиданты (α-, β-, γ- и δ-токоферолы, аскорбиновую кислоту, β- каротин, мочевую кислоту, убихинон), а также ферменты антиоксидантной защиты (супероксиддисмутазу, каталазу, глутатионпероксидазу, глутатионредуктазу, церрулоплазмин). С целью повышения эффективности антиоксидантной защиты клеток организма в пищевые продукты часто добавляют искусственные антиоксиданты – пропилгаллат, бутилированный гидроксианизол и бутилированный гидрокситолуол. Антиоксиданты способны терминировать процесс ПОЛ или нейтрализовывать продукты ПОЛ. В связи с этим антиоксиданты подразделяют на два класса:
1)превентивные антиоксиданты, снижающие скорости инициации цепных реакций;
2)гасящие (прерывающие цепь) антиоксиданты, препятствующие развитию цепных реакций.
Кпервому классу антиоксидантов относят каталазу и другие пероксидазы, способные разрушать гидроперекиси (ROOH), а также агенты, образующие хелатные комплексы с металлами – диэтилентриаминпентаацетат и этилендиаминтетраацетат. В качестве антиоксидантов второго класса, пре-
29
рывающих цепные реакции, часто выступают фенолы или ароматические амины. В условиях in vivo главными антиоксидантами второго класса являются супероксиддисмутаза, которая улавливает в водной фазе свободные супероксидные радикалы, а также витамин Е, улавливающий свободные ROO·- радикалы в липидной фазе.
Недостаток антиоксидантов в организме приводит к инициированию процессов непосредственного присоединения кислорода к окисляемым соединениям с образованием альдегидов, пероксидов, свободных радикалов, которые в химическом отношении являются высокотоксичными и весьма агрессивными веществами. В этом случае нарушается нормальное течение физиологических процессов: блокируется депонирование энергии в форме АТР, энергия рассеивается в виде тепла. Повреждаются клеточные мембраны, ферменты, структурные белки. Повреждение структурных белков, в частности, проявляется в нарушении синтеза эластических волокон артериальных стенок, что служит причиной предрасположенности к развитию атеросклероза. Этому явлению способствует состояние гиперхолестеринемии, возникающее за счет нарушения процессов превращения стеринов в желчные кислоты в присутствии пероксидов липидов, которые в норме осуществляются в гепатоцитах.
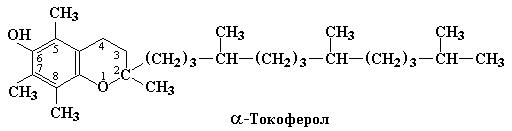
Одним из высокоэффективных компонентов системы антиоксидантной защиты является витамин Е, который представлен смесью из 4 токоферолов и 4 токотриенолов. На рис. 3.1 приведена структурная формула α- токоферола.
Рис. 3.1 Структурная формула α-токоферола. У β- и γ-токоферолов в положениях 7 и 5, соответственно, заместителем является Н. δ-токоферол содержит Н в 5 и 7 положениях.
Как витамины наиболее активны α-, β- и γ-токоферолы, эффективность которых выражается отношением 100:40:8. С другой стороны, наиболее сильные антиоксидантные свойства присущи δ-токоферолу.
30
Витамин Е является первым уровнем защиты клеточных и субклеточных мембранных фосфолипидов от перекисного окисления. Токоферолы действуют как антиоксиданты, прерывающие цепи окисления, благодаря их способности переносить фенольный водород на пероксидный радикал (рис. 3.2). Феноксирадикал токоферола является резонансно-стабилизированной и достаточно устойчивой структурой.
Рис. 3.2 Гасящее цепную реакцию антиоксидантное действие токоферолов по отношению к ROO·-радикалам.
Исходя из сказанного выше, следует, что токоферолы практически не вовлекаются в процесс цепной реакции. В ходе окисления хроманолового кольца и боковой цепи токоферолов образуется продукт, не являющийся свободным радикалом:
Этот продукт образует коньюгат с глюкуроновой кислотой и экскретируется с желчью. Необходимо отметить, что в подобные реакции с ROO·- радикалами вступают фенолы, причем феноксирадикалы фенолов отличаются повышенной устойчивостью, особенно замещенные по 2-ому, 4-ому или 6-
ому положениям.
31
Следовательно, способность к образованию относительно устойчивых свободных радикалов токоферолов можно объяснить как наличием хроманольной структуры, так и присутствием метильных заместителей в положениях 5 и 7 (см. рис. 3.1).
Витамин Е выполняет в организме млекопитающих, по крайней мере, две метаболические функции. Во-первых, он служит сильнодействующим природным жирорастворимым антиоксидантом и, во-вторых, выполняет специфическую роль в метаболизме селена. Действительно, многочисленные данные свидетельствуют о наличии взаимосвязи между пищевой потребностью в селене и витамине Е. Поскольку недостаточность витамина Е сопровождается низким содержанием селенида (Se2-), было высказано предположение, что витамин Е предохраняет восстановленный селен от окисления с образованием селенита (SeO32-) или других более окисленных состояний селена.
Антиоксидантное действие α-токоферола сохраняется при высоких концентрациях кислорода, поэтому неудивительно, что витамин Е накапливается в богатых липидами областях, контактирующих со средой, где поддерживается высокое парциальное давление кислорода - в мембранах эритроцитов и клеток дыхательных путей. Типичное проявление нехватки токоферолов – выраженная миодистрофия, которая охватывает как скелетные (особенно, диафрагмальную), так и гладкие мышцы, а также миокард. Происходит деструкция митохондрий, фрагментация мышечных волокон, появляются микронекрозы. Гиповитаминоз Е сопровождается тканевой гипоксией в органах с высокой потребностью в кислороде. Известно снижение резистентности эритроцитов к гемолизу, наступающее при гиповитаминозе Е. Вследствие перекисной трансформации эритроцитарных мембран, красные кровяные клетки приобретают шиповатую форму (акантоцитоз). У недоношенных детей, а также у обезьян может проявляться острая гемолитическая анемия, купируемая витамином Е. Таким образом, действие витамина Е и селена состоит, по-видимому, в предохранении клеточных и субклеточных компонентов от повреждения перекисями, обеспечивая их целостность.
Однако даже в присутствии адекватного количества витамина Е может происходить образование некоторого количества пероксидов. Вторым уровнем защиты мембран от разрушающего действия перекисей служит селенопротеид – глутатионпероксидаза – катализирующий реакцию окисления глутатиона пероксидом водорода.
Принимая во внимание все сказанное выше можно утверждать что, определение степени насыщенности организма антиоксидантами является важным прогностическим и диагностическим показателем для характеристики состояния здоровья и болезни организма.
32