27 Структура и функции НК. Особенности строения и роль различных видов ДНК (ядерная, митохондриальная, рибосомальная, сателлитная и др.). Особенности структуры ДНК вирусов и фагов. Нуклеиновые кислоты - это полинуклеотиды
Особенности нуклеиновых кислот
|
Признак |
ДНК |
РНК |
|
Месторасположение в клетке |
Ядро, небольшое количество в цитозоле |
Цитоплазма, небольшое количество в ядре |
|
Сахар |
дезоксирибоза |
рибоза |
|
Главные основания |
аденин, гуанин, цитозин, тимин |
аденин, гуанин, цитозин, тимин |
|
Молекулярная масса |
>109 |
20000-109 |
|
Функция |
Хранение и передача генетической информации |
Синтез белков |
|
Форма вторичной структуры молекулы |
Двойная спираль |
Одноцепочечная молекула |
Нуклеиновые кислоты представляют собой полинуклеотиды. Нуклеотиды построены из трех компонентов: пиримидинового или пуринового основания, пентозы и фосфорной кислоты. Нуклеотиды связаны между собой в цепь фосфодиэфирной связью. Она образуется за счет этерификации ОН - группы С-З’ пентозы одного нуклеотида и ОН - группы фосфатного остатка другого нуклеотида. В результате один из концов полинуклеотидной цепи заканчивается свободным фосфатом (Р-конец или 5’-конец). На другом конце цепи имеется неэтерифицированная ОН - группа у С-З’ пентозы (З’ - конец)
Первичнаяструктура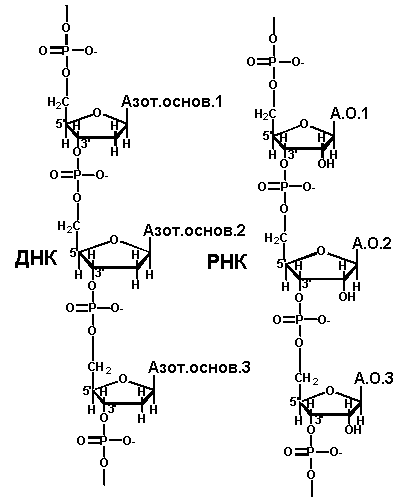
Вторичная структура
Молекула ДНК представляет собой правозакрученную спираль, состоящую из двух полинуклеотидных цепей с антипараллельным ходом. Это означает, что 3’-концу одной цепи соответствует 5’-конец другой цепи и наоборот.
Остатки оснований направлены внутрь спирали. На один виток спирали приходится 10 пар оснований. Цепи ДНК не идентичны, так как нуклеотидный состав их различен, однако первичная структура одной цепи предопределяет нуклеотидную последовательность другой цепи, то есть они комплементарны друг другу. Это связано с существованием комплементарных пар оснований.
Функции нуклеиновых кислот заключаются
в хранении,
реализации
и передаче генетической информации.
Ядерная ДНК
В препарате хроматина на долю ДНК приходится обычно 30-40%. Эта ДНК представляет собой двухцепочечную спиральную молекулу.
ДНК эукариотических хромосом представляет собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. Тем самым в составе генома человека должно встречаться более 50 000 репликонов, участков ДНК, которые синтезируются как независимые единицы. Эти репликоны имеют начальную и терминальную точки синтеза ДНК.
ДНК эукариотических клеток гетерогенна по составу, содержит несколько классов последовательностей нуклеотидов:
часто повторяющиеся последовательности (>106 раз), входящие во фракцию сателитной ДНК и не транскрибирующиеся;
фракция умеренно повторяющихся последовательностей (102 -105), представляющих блоки истинных генов, а также короткие последовательности, разбросанные по всему геному;
фракция уникальных последовательностей, несущая информацию для большинства белков клетки.
Митохондриальная ДНК (мтДНК) — ДНК, локализованная (в отличие отядернойДНК) вмитохондриях. У большинства многоклеточных организмов митохондриальный геном наследуется по материнской линии и состоит изкольцевой ДНК размером 16600 нуклеотидов. Геном митохондрий человека кодирует следующие белки и РНК:
|
NADH-дегидрогеназа (комплекс I) |
|
Кофермент Q - цитохром c редуктаза/Цитохром b (комплекс III) |
|
цитохром c оксидаза (комплекс IV) |
|
АТФ-синтетаза |
|
рРНК |
|
тРНК |
Многие митохондриальные белки кодируются генами ядерного генома, синтезируются в цитоплазме и затем транспортируются в митохондрии.
Рибосомальная ДНК
Фракция умеренно повторяющихся (от 102 до 105 раз) последовательностей принадлежит к большому классу участков ДНК, играющих важную роль в обменных процессах. В эту фракцию входят гены рибосомных ДНК, многократно повторенные участки для синтеза всех тРНК.
Сателитная ДНК На долю повторяющихся последовательностей приходится более половины генома человека. Эти последовательности ДНК чаще всего состоят из участков длиной 5-500 пар, расположенных один за другим (тандемно), обычно не транскрибируются и часто располагаются в центромерах хромосом. Такие последовательности обозначаются как спутниковая или сателитная ДНК. Сателлитная ДНК, или фракция ДНК с часто повторяющимися последовательностями, может участвовать в узнавании гомологичных районов хромосом при мейозе. По другим предположениям, эти участки играют роль разделителей (спейсеров) между различными функциональными единицами хромосомной ДНК.
ДНК вирусов и фагов
Геном вирусов содержит ДНК либо РНК. ДНК вирусов может быть двуцепочечная (двунитевая) или одноцепочечная (однонитевая), с последующим увеличением до двуцепочечной в клетках хозяина. ДНК имеет линейную форму или кольцевую.
Спустя 25 лет после открытия вируса, канадский ученый Феликс Д'Эрел открыл новую группу вирусов, поражающих бактерии. Они так и были названы бактериофагами (или просто фагами).
Жизненный цикл бактериофагов.
Фаг приближается к бактерии, и хвостовые нити связываются с рецепторными участками на поверхности бактериальной клетки.
Хвостовые нити изгибаются и "заякоривают" шипы и базальную пластинку на поверхность клетки; хвостовой чехол сокращается, зачтавляя полый стержень входить в клетку; этому способствует фермент - лизоцим, который находится в базальной пластинке; таким образом ДНК вводится внутрь клетки.
ДНК фага кодирует синтез ферментов фага, используя для этого белоксинтезирующий аппарат (рибосомы и т.п.) хозяина.
Фаг тем или иным способом инактивирует ДНК хозяина, а фермент фага совсем расщепляет ее; ДНК фага починяет себе клеточный аппарат
ДНК фага реплицируется и кодирует синтез новых белков.
Новые частицы фага, образующиеся в результате спонтанной самосборки белковой оболочки вокруг фаговой ДНК; под контролем ДНК фагов синтезируется лизоцим.
Лизис клетки, т.е. клетка лопается под воздействием лизоцима; высвобождается около 200-1000 новых фагов; фаги индуцируют другие клетки.
Жизненный цикл фага составляет 30 минут.
28. Особенности строения и роль различных видов рнк - информационной, рибосомальной, транспортной, вирусной.
В зависимости от локализации и функции различают несколько типов РНК:
Около 80% всей клеточной РНК находится в цитозоле в составе рибосом. Это рибосомная РНК. Главная функция этой РНК - структурная организация рибосомы, места, где происходит сборка полипептидной цепи.
Еще 2 типа РНК выполняют специальные функции в синтезе белка.
15% РНК клетки носит название транспортной РНК и выполняет 2 основные фунции: активирование и перенос аминокислот и адапторную функцию. Последняя позволяет перевести "язык" генетического кода (последовательность нуклеотидов) на "язык" структуры белковой молекулы (аминокислотная последовательность). Каждая аминокислота имеет несколько специфичных для нее транспортных РНК.
 А
А
Структура транспортной РНК. Вторичная структура (А) напоминает лист клевера. В полинуклеотидной цепи тРНК часто встречаются необычные нуклеотиды (дигидроуридиловый, псевдоуридиловый, риботимидиловый), отсюда и название некоторых структурных элементов. Пространственная (третичная) структура тРНК напоминает букву Г или латинскую перевернутую букву L
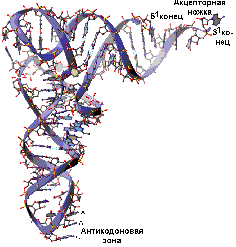 В
В
Около 5 % всей РНК клетки это информационная РНК. Эта РНК является комплементарной копией генов, кодирующих белки. Она используется как матрица на рибосоме во время сборки полипептидной цепи.
Принцип организации первичной структуры РНК тот же, что и у ДНК. На уровне вторичной структуры молекула РНК формирует однонитиевые молекулы, отдельные участки молекулы, комплементарные друг другу, образуют двунитевые "шпильки", стабилизируемые также как и двойная спираль ДНК. В формировании третичной структуры РНК важную роль играют белки. При этом возникают большие внутриклеточные частицы: рибосомы, информосомы.
Исключение составляет небольшая по размерам транспортная РНК (70-80 мононуклеотидов). Ее стабильность и устойчивость к действию гидролизующих ферментов объясняется высоким содержанием в ней минорных азотистых оснований.
Отдельно следует упомянуть вирусную РНК. У некоторых типов вирусов РНК играет роль хранительницы генетической информации. Проникновение такого вируса в клетки требует предврительного синтеза ДНК по матрице РНК этого вируса. В дальнейшем вирус ведет себя как и вирус, имеющий ДНК.
|
Виды РНК |
Размер в нуклеотидах |
|
gРНК - геномные РНК |
10000-100000 |
|
mРНК - информационные (матричные) РНК |
100-100000 |
|
tPHK - транспортные РНК |
70-90 |
|
rРНК - рибосомные РНК |
несколько дискретных классов от 100 до 500000 |
|
sРНК - малые РНК |
100-300 |
29. Механизмы хранения и передачи наследственной информации - репарация, репликация (строение репликативной вилки), транскрипция, трансляция, характеристика основных ферментов и кофакторов.
Репликация («самоудвоение») - перенос генетической информации в пределах одного класса нуклеиновых кислот, т.е. от ДНК к ДНК (происходит полное копирование информации).
Транскрипция («переписывание») – перенос информации между разными классами нуклеиновых кислот: от ДНК к РНК (от РНК к ДНК – у вирусов), т.е. происходит копирование отдельных участков. В ходе транскрипции образуются все виды РНК (м-РНК, т-РНК, р-РНК)
Трансляция – перенос генетической информации от м-РНК к белку, т.е. в пределах разных классов молекул: перевод информации с «языка» нуклеиновых кислот на «язык» полипептидной цепи.
Репарация ДНК – ограниченная репликация, исправление поврежденных участков ДНК ДНКазами, затем ДНК-полимеразы заполняют пробел и концы «сшиваются» ДНК лигазами (см. рисунок).
Репликация ДНК (воспроизведение генотипа) происходит по полуконсервативному механизму, то есть дочерняя цепь ДНК синтезируется на материнской цепи, называемой обычно матрицей. Следовательно, вновь образованные двухспиральные молекулы состоят из одной «новой» и одной «старой» цепи
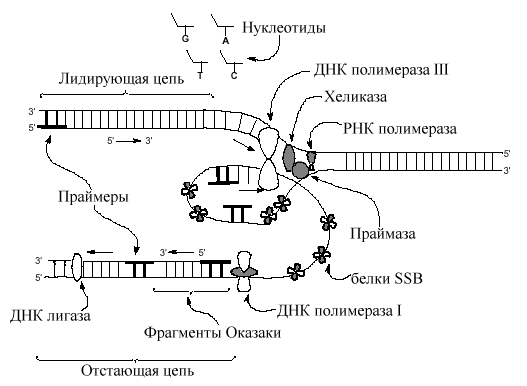
1. Все ДНК-полимеразы нуждаются для начала своей работы в предварительно синтезированных олигонуклеотидах – затравках, 3’-ОН группа которых используется для удлинения цепи. (Эти короткие отрезки, обычно, рибонуклеотидов синтезируются специальными РНК-полимеразами. По названию затравки – праймер эти полимеразы получили название праймаз). Длина праймеров составляет 10-12 нуклеотидов. Праймеры синтезируются на обеих цепях.
2. Далее праймер достраивается ДНК-полимеразой – III. Т.к. все ДНК-полимеразы формируют дочерние цепи в одном направлении 5’ 3’, то одна цепь синтезируется непрерывно (лидирующая, ведущая цепь), а другая - прерывисто (отстающая). ДНК-полимераза формирует димер, связанный с другими необходимыми в репликативной вилке белками (реплисома). Одна из цепей матричной ДНК (отстающая), временно образует петли вокруг реплисомы так, что димер ДНК-полимеразы получает возможность перемещаться по обеим цепям в одном 3' 5 ' направлении одновременно. Короткие (до 1000 пар нуклеотидов) отрезки ДНК, образующиеся на отстающей цепи, получили название фрагментов Оказаки по имени Ф. Оказаки, впервые (1968) указавшего на прерывистый характер синтеза ДНК. 3.Так как репликативная вилка довольно быстро (1000 нулеотидов в сек) продвигается по матрице, вновь синтезируемые дочерние цепи и родительские цепи матрицы сразу формируют двойные спирали ДНК. Предполагается, что только маленький отрезок матричной двойной спирали находится в одноцепочечном состоянии в данный отрезок времени.
3. Праймеры ведущей и отстающей цепей (10-12 пар нуклеотидов) удаляются ДНК полимеразой I, обладающей репарирующей функцией с одновременной заменой рибонуклеотидов дезоксирибонуклеотидами.
4. Промежутки, которые возникают между 3 '-ОН и 5 '- фосфатом, «сшиваются» лигазами ДНК, завершая тем самым процесс репликации
транскриптон (оперон) - единица транскрипции.
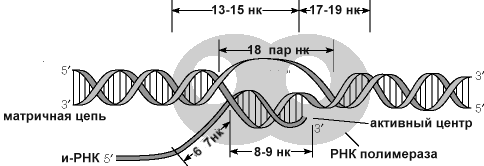
Синтез молекул РНК начинается в определенных местах ДНК, называемых промоторами и завершается в терминаторах. Участок ДНК, ограниченный промотором и терминатором, представляет собой единицу транскрипции - транскриптон (оперон у прокариот). В пределах каждого транскриптона копируется только одна из двух нитей ДНК, которая называется значащей или матричной. Во всех транскриптонах, считываемых в одном направлении, значащей является одна нить ДНК, в транскриптонах, считываемых в противоположном направлении, значащей является другая нить ДНК.
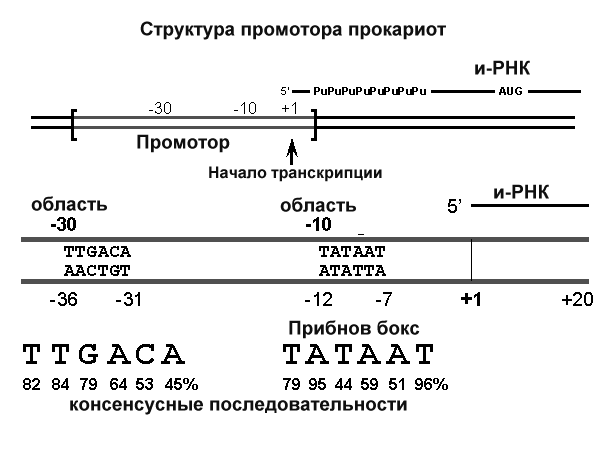
Разбиение ДНК на множество транскриптонов обеспечивает возможность независимого считывания разных генов, их индивидуального включения и выключения.
2 коротких консервативных участка последовательностей, расположенных на расстоянии 35 нуклеотидных пар в сторону 5’ конца от начала транскрипции (область -30 на рис.) и на расстоянии 10 нуклеотидов в ту же сторону (область -10). АТ богатый участок области -10 получил название ТАТА бокса или Прибнов бокса. Считается, что эта последовательность может быть легко денатурирована РНК-полимеразой и является удобным местом начала транскрипции.
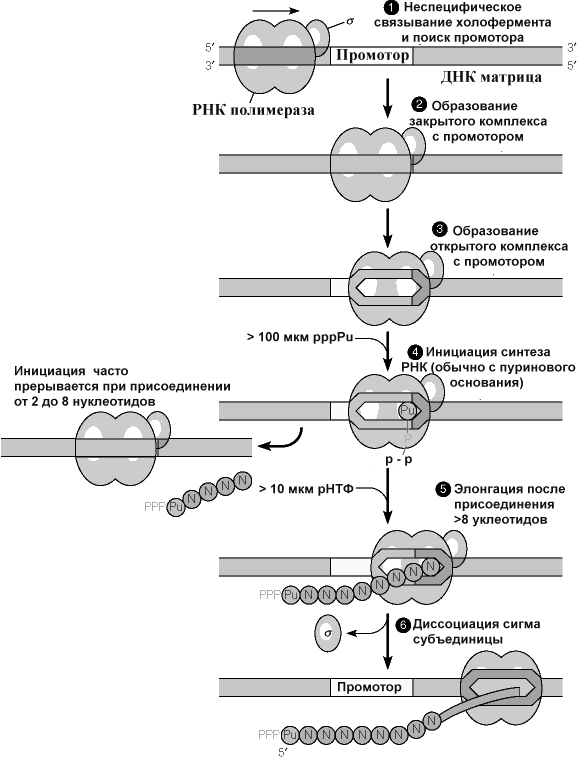
Первый шаг в транскрипции - связывание РНК-полимеразы с ДНК, с последующим перемещением к инициирующему участку ДНК. Поиск промотора РНК-полимеразой начинается (на рисунке этап 1) с неспецифического связывания фермента с ДНК, и последующего перемещения по ДНК, без диссоциации до достижения последовательности.
промотора, с которым фермент связывается со значительно более высоким сродством. Этому способствует фактор, потому что базовый фермент имеет одинаковое сродство к любой последовательности ДНК.
Первоначальное взаимодействие РНК-полимеразы и промотора образует закрытый комплекс, цепь ДНК в этом комплексе не раскручена.
РНК-полимераза раскручивает несколько пар оснований ДНК, от приблизительно -10 до -1, образуя открытый промоторный комплекс, названный так потому, что в нем цепи ДНК открыты, или раскручены, фермент готов начать синтез цепи РНК.
После синтеза первых 10 нуклеотидов, субъединица отделяется от комплекса транскрипции, и дальнейший процесс транскрипции катализируется базовой (коровой) полимеразой (см. рисунок, шаги 5 и 6). Хотя и субъединица покинула фермент, элонгирующий комплекс является довольно устойчивым. Элонгация, таким образом, продолжается до заключительного этапа. Во время элонгации (см. рисунок выше, шаги 5 и 6), коровый феремент перемещается по двуцепочечной матрице ДНК, одновременно раскручивает ДНК, копируя дноцепочечную матрицу ДНК с образованием транскрипта (вновь синтезируемая РНК). Фермент скручивает матрицу позади 3 ' конца растущей цепи РНК
В областях окончания транскрипции происходит диссоциации РНК-полимеразы и ДНК