

Объекты и методы
Объектами исследования являлись 28-летние опытные культуры ели европейской плантационного типа, созданные на дренированной легкосуглинистой почве в кисличном типе леса (эдатоп С2) Псковского лесничества (кв.35)с густотой посадки 1000 и 4000 экз./ га.Подготовка почвы произведена методом нарезки плужных борозд плугом ПЛО-400. Посадка проведена в пласт 4-летними саженцами, выращенными из семян производственного сбора. В культурах до 15-летнего возраста проведено 3 агротехнических и 3 лесоводственных ухода со сплошной вырубкой лиственных пород. Поэтому на индивидуальный характер роста деревьев оказывали влияние внутривидовые отношения, а межвидовые – отсутствовали. На период обследования число растущих деревьев ели в редких и густых культурах составило, соответственно, 936 и 3280 экз./га. Различие по запасу стволовой древесины оказалось незначительным: 291 м3/га в редких и 305 м3/га в густых культурах. Для анализа роста и феногенетической структуры было изучено всего 185 деревьев ели, в том числе 99 экз. в редких посадках и 86 экз. – в густых. У каждого дерева определялись следующие показатели: высота в 5, 10 и 28 лет, ежегодные (от 5 до 10-летнего возраста) линейные приросты, диаметр у основания ствола и на высоте 1.3 м, диссимметрия (левизна-правизна) филлотаксиса осевого побега. При изучении диссимметрической структуры кроны у каждого дерева обследовалось 30–40 побеговIпорядка ветвления. Определение левых (L) и правых (D) форм побегов проводилось по методике А.В. Хохрина [1977]. Для электрофоретического анализа изоферментов использованы вегетативные почки; у каждого дерева было изучено 26 аллозимных локусов:Fdh,Fe-1,Fe-2,Gdh,Got-1, Got-2, Got-3, Idh-1, Idh-2, Lap-1, Lap-2, Mdh-1a, Mdh-1, Mdh-2, Mdh-3,Ndh-2,Pepca,Pgd-2,Pgd-3,Pgi-2,Pgm-1,Pgm-2,Skdh-1,Skdh-2,Sod-1,Sod-2. Данный анализ выполнен в Институте общей генетики им. Н.И. Вавилова РАН.
Результаты и обсуждение
Наиболее высокий отпад деревьев ели на момент обследования произошел в культурах с высокой густотой посадки и составил 18% от числа высаженных растений, тогда как в редких посадках отпад составил 6.4%.
Данные табл. 1 показывают, что в густых культурахпроцессы дифференциации и самоизреживания ускорены и в них наблюдается наибольшая вариабельность высоты (СV= 16.2%), диаметра (СV= 29.7%) и объема ствола (СV= 61.4%). В густых культурахD-формы ели превосходилиL-формы по средним показателям высоты на 10.7%, по диаметру на 18.6% и по объему ствола на 32.7%. Левые формы отличались здесь наибольшей вариабельностью: коэффициент изменчивостиL-форм по диаметру составил 30.7%, по высоте – 17.8% и объему ствола 72.5%).
В культурах с редкой
посадкой, наоборот,L-формы
ели имели более высокие показатели: по
диаметру на 11.6%, по высоте на 7.7% и объему
ствола на 23.5%.D-формы
здесь обладали слабым ростом и отличались
отL-форм наибольшим
уровнем дифференциации. Выявленные
различия по размерам уLиD-форм ели в редких и
густых культурах статистически достоверны
(t![]() =
2.6-4.9).
=
2.6-4.9).
Приведенные показатели свидетельствуют, что на повышение популяционной плотности левые энантиоморфы реагируют замедлением роста и увеличением изменчивости размеров стволов, тогда как правые, напротив, ускоряют темпы роста и снижают дифференциацию. Вполне очевидно, что LиD-энантиоморфы ели обладают противоположными онтогенетическими реакциями на эффекты загущения: при низкой плотности популяции адаптивные преимущества имеютL-формы, а при высокой –D-формы. Адаптивная разнокачественность энантиоморф ели, вероятно, связана со световой конкуренцией. Ранее было установлено, чтоL-формы сосны и ели отличаются большим светолюбием, аD-формы, наоборот, обладают большей теневыносливостью [Хохрин, 1977; Голиков, 1985;Golikov, 1989].Поэтому левые формы как наиболее светолюбивые имеют преимущества в разреженном древостое, а правые формы как более теневыносливые, наоборот, лучше приспособлены к росту в густых ценозах. Можно предположить, что L-формы лучше используют прямой солнечный свет, а D-формы – рассеянный. Не исключено, что L-формы ели в редких посадках способны максимально использовать жизненное пространство, тогда как D-формы ели обладают более высоким синтезом органического вещества на единицу площади в густых культурах.
Таблица 1
Влияние густоты посадки на изменчивость роста 28-летних энантиоморф ели
|
Энантио-морфа |
Изучено деревьев, экз. |
Диаметр (D1.3), см |
Высота, м | ||||
|
|
СV,% |
t |
|
СV,% |
t | ||
|
Густота
посадки 1000 экз. га | |||||||
|
L |
52 |
24.1 0.3 |
10.2 |
4.3 |
15.4 0.1 |
6.3 |
4.9 |
|
D |
47 |
21.6 0.5 |
17.2 |
14.3 0.2 |
11.1 | ||
|
L+D |
99 |
22.9 0.3 |
15.1 |
|
14.9 0.1 |
9.6 |
|
|
Густота
посадки 4000 экз. га | |||||||
|
L |
38 |
11.3 0.6 |
30.7 |
2.7 |
12.2 0.4 |
17.8 |
2.6 |
|
D |
48 |
13.4 0.5 |
26.8 |
13.5 0.3 |
13.9 | ||
|
L+D |
86 |
12.5 0.4 |
29.7 |
|
12.9 0.2 |
16.2 |
|
Для оценки влияния густоты посадки на процессы самоизреживания и самоорганизации феногенетической структуры модельных популяций у энантиоморф ели по стабильности их роста в онтогенезе (5, 10 и 28 лет) выделено 4 типа развития:
I. Стабильно быстрый рост (L+,D+) – высота энантиоморф выше среднепопуляционной на всех возрастных этапах роста;
II. Стабильно медленный рост (L– ,D–) – высота энантиоморф ниже средней на всех возрастных этапах роста;
III. Ускоренный (L– +,D– +) – высота в 5-10 лет ниже средней, в дальнейшем – выше;
IV. Замедленный (L+ – ,D+ –) – высота в 5-10 лет выше средней, в дальнейшем – ниже.
Показатели роста моделей по этим 4 типам развития деревьев представлены в табл. 2.
Из всех типов роста наибольший интерес представляют генотипы со стабильно быстрым ростом.Их диссимметрический состав зависит от густоты древостоя:в редких культурах среди всех типов роста явно доминируют L+ формы (31.3%), а в густых, напротив, чаще встречаются формы D+ (31.4%).Зеркально обратная закономерность по частоте встречаемости проявляется у энантиоморф со стабильно медленным ростом. Так, доляL– форм существенно возрастает в густых культурах и составляет 18.6%, а в редких – уменьшается до 9.1%. В густых культурах, напротив,D– формы ели встречаются меньше (10.5%), а в редких – в 2 раза чаще (20.2%). Число деревьев с ускоренным (L– +,D– +) и замедленным ростом в редких культурах оказалось равным – по 14.1%, т.е. их ранговое перемещение взаимно уравновешивается. В густых культурах данное равновесие нарушено в пользу энантиоморф с ускоренным типом роста, где они оказались более устойчивыми, чем энантиоморфы с замедленным ростом
В редких культурах большую часть запасов древесины (59,5%) накапливают в своих стволах левые формы, а в густых – правые формы (65.2%), при отмеченном выше совсем небольшом численном перевесе этих форм, соотвественно, в редких и в густых культурах (рис. 1). Особенно примечательно явное доминирование запасов древесины в стволах стабильно быстрорастущих левых (L+) и правых (D+) форм, формирующих особенно крупные стволы; именно эти генотипы в ценозе являются центрами накопления запасов древесины, и именно на них должны быть направлены усилия селекционеров и лесоводов (рис. 2).
Изложенное выше позволяет полагать, что плотность популяции оказывает существенное влияние на диссимметрический состав деревьев ели и на состав типов их роста и численное соотношение энантиоморф можно использовать в качестве важной характеристики, указывающей направление естественного отбора и микроэволюционных процессов.
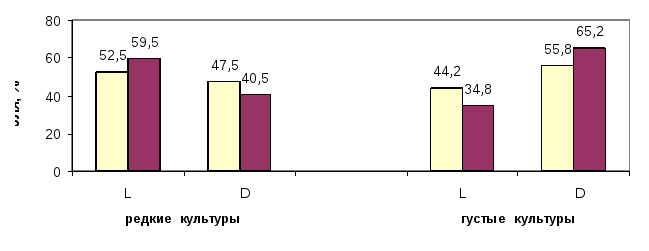
Рис. 1. Левые (L) и правые (D) формы ели в редких и густых культурах и их доли по числу деревьев (столбик слева) и по запасу древесины (столбик справа)
|
|
|
|
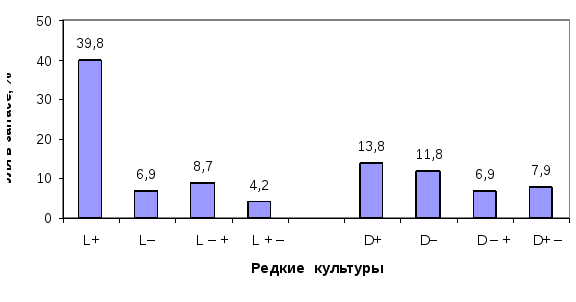
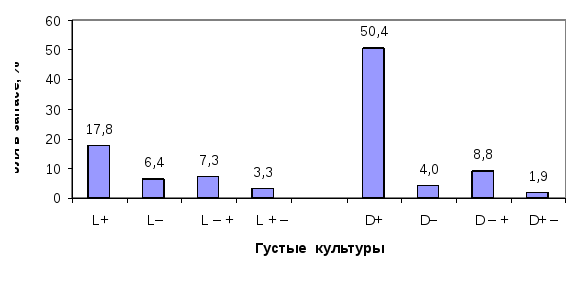
Рис. 2. Доли в запасах древесины, приходящиеся на энантиоморфы с разным типом роста в редких и в густых культурах
Проведенный анализ показал, что тип роста деревьев ели в редких и густых культурах тесно связан с диссимметрическими признаками и реакция LиD-форм на изменение плотности ценоза зеркально противоположная. Эта особенность, очевидно, является основой для функционирования и адаптации того или иного генотипа к плотности древостоя. На популяционном уровне данный механизм действует по кибернетическому принципу положительной и отрицательной обратной связи. Можно предположить, что быстрота роста деревьев ели, растущих в условиях высокой и низкой конкуренции, обуславливаются различными генетическими системами, контролирующих фото-ассимилирующий аппарат. Доказательством этого является тот факт, что светолюбивые левые формы имеют преимущество в редких культурах, а теневыносливые правые – в густых культурах.
Библиографический список
Авдеев Э.А., Голиков А.М. Влияние уровня гетерозиготности на репродуктивную и наследственную не- равноценность плюсовых деревьев ели европейской // Современное состояние, проблемы и перспективы лесовосстановления и лесоразведения на генетико-селекционной основе. Материалы международной науч.-практ. конф. Гомель: Ин-тут леса НАН Беларуси. 2009. С.19-23.
Алтухов Ю.П. Генетические процессы в популяциях. М.: Наука, 1989. 328 с.
Алтухов Ю.П. Аллозимная гетерозиготность, возраст первой репродукции и продолжительность жизни у древесных // Докл. РАН. 1996. Т. 351. № 6. С. 837-840.
Алтухов Ю.П., Корочкин Л.И., Рычков Ю.Г. Наследственное биохимическое разнообразие в процессах эволюции и индивидуального развития // Генетика.1996. Т.32. № 11. С.1450-1473.
Бакшаева В.И. К вопросу о явлении диссимметрии у сосны обыкновенной в Карелии // Повышение продуктивности хвойных лесов Карелии. Петрозаводск. Карельский ФАН СССР. 1975. Т. 22. Вып. 3. С. 74-106.
Глазко В.И., Созинов И.А. Генетика изоферментов животных и растений. Киев: Урожай, 1993. 528 с.
Голиков А.М. Формы сосны обыкновенной и их селекционное значение в условиях Псковской области: Автореф. дис. … канд. с.–х. наук: 06.03.01.Свердловск, 1985. 18 с.
Голиков А.М. Влияние экологических факторов на наследственную неравноценность диссимметричных форм плюсовых деревьев сосны обыкновенной // Труды СПбНИИЛХ. СПб, 2004. Вып. 2(12). С. 117-131.
Голиков А.М. Рост и формовая структура потомства ели европейской в зависимости от гетерозиготности деревьев и условий произрастания // Лесоведение. 2007. № 4. С.51-58.
Голиков А.М. Диссимметрическая и генетическая разнокачественность фенологических форм плюсовых деревьев ели европейской // Лесоведение. 2007. № 1. С. 49-56.
Голиков А.М. Адаптивная и генетическая разнокачественность энантиоморф плюсовых деревьев ели европейской на Северо-Западе России // Современное состояние, проблемы и перспективы лесовосстановления и лесоразведения на генетико-селекционной основе. Материалы междунар. науч.-практ. конф. Гомель. Ин-т леса НАН Беларуси. 2009. С. 33-36.
Голиков А.М. Теоретическое и прикладное значение эколого-диссимметрийного подхода в исследовании формовой и генетической структуры популяций видов хвойных // Наука о лесе ХХI века. Материалы междун. науч.-практ. конф., посвященной 80-летию Ин-та леса НАН Белоруси. Гомель: Ин-т леса НАН Белоруси. 2010. С.157-160.
Голиков А.М. Диссимметрическая и генетическая дифференциация плюсовых деревьев сосны обыкновенной в разных типах леса // Лесоведение. 2011. № 3. С. 63-72.
Голиков А.М. Эколого-диссимметрический и изоферментный анализ формовой и генетической структуры модельных популяций сосны обыкновенной // Лесоведение. 2011. № 5. С. 46-53.
Голиков А.М. Использование эколого-диссимметрийного подхода для интенсификации процесса генетического улучшения хвойных лесов // Инновации и технологии в лесном хозяйстве. Материалы междун. науч.-практ. конф. Санкт-Петербург: Тр.ФГУ «СПбНИИЛХ», вып.1(24), ч.1. 2011. С. 63-67.
Долотовский И.М., Драгавцев В.А. Влияние взаимодействия генотипа с условиями популяционной плотности и года на проявление количественных признаков у гороха посевного // Генетика. Т. ХХIV. № 6. 1988. С. 1057-1064.
Маслаков Е.Л. Формирование сосновых молодняков // М.:Лесн.пром-сть. 1984. 168 с.
Роне В.М. Генетический анализ лесных популяций // М.: Наука, 1980. 160 с.
Рогозин М.В., Разин Г.С. Лесные культуры Теплоуховых в имении Строгановых на Урале: история, законы развития, селекция ели. Пермь, 2011. 192 с.
Сукачев В.Н. О влиянии интенсивности борьбы между растениями на их развитие // ДАН СССР. 1941. Т.20. № 8. С. 752-755.
Сукачев В.Н. Избранные труды. Т.1 Основы лесной типологии // Л. Наука. 1972. 418 с.
Хохрин А.В. Внутривидовая диссимметрическая изменчивость древесных растений в связи с их экологией: Автореф. дис. … д–ра. биол. наук. 03.00.16. Свердловск, 1977. 49 с.
Golikov A.M. Adaptive disparity of dissymmetrical forms of Pinus silvestris L. and Picea abies L. Karst. in the north–west of Russian SFSR. // Symmetry of structure. Interdisciplinary symmetry symposia, 1. Budapest: The Hungarian Academy of Sciences, 1989. P. 168-171.