


ХОЛЕСТЕРИН
Желательный уровень - менее 5,2 ммоль/л
Пограничный уровень - 5,2-6,2 ммоль/л
Критический уровень - более 5,2 ммоль/л

Окисление жирных кислот
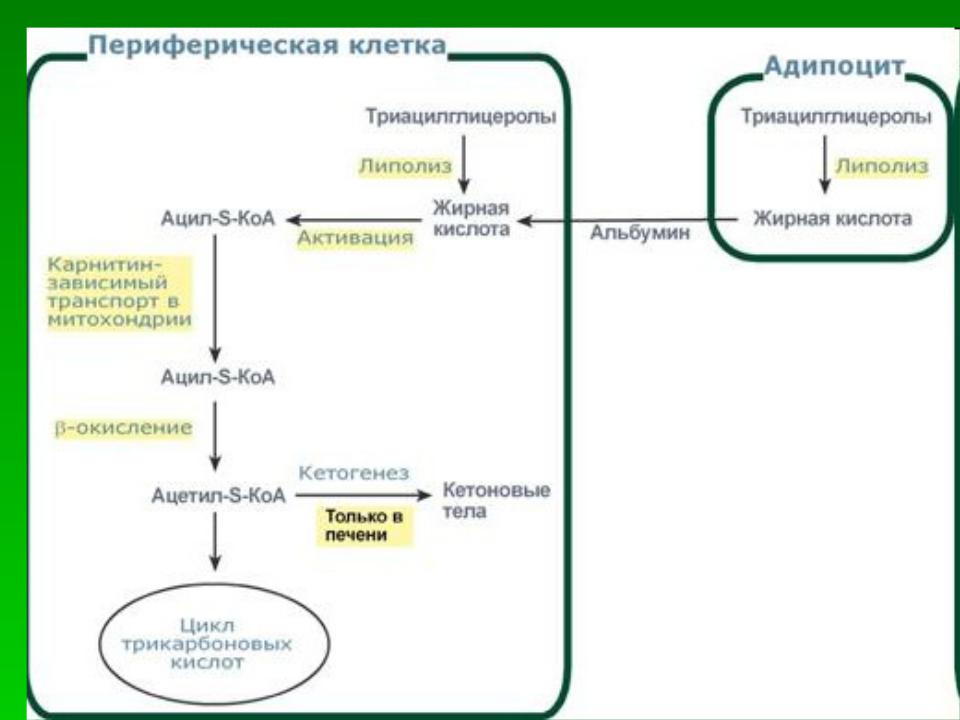
Жирные кислоты во многих тканях (в первую очередь в скелетных мышцах и миокарде) вовлекаются в специфический процесс – b-окисление, протекающее в митохондриях. Все превращения ЖК начинаются с их активации — образования ацил-КоА.
Мембрана митохондрий непроницаема для жирных кислот, в том числе и в форме ацил-КоА. Их перенос обеспечивается карнитином.
В митохондриях на митохондриальном матриксе происходит окисление жирных кислот, сопряженное с окислительным фосфорилированием (с синтезом АТФ). b- окисление протекает по следующей схеме:
В первой реакции происходит дегидрирование при участии ФАД-зависимой дегидрогеназы. Затем присоединение воды (гидроксил – в b-положение) с образованием b-гидроксиацил-КоА. Дегидрирование НАД-зависимой дегидрогеназой у b-углеродного атома приводит к образованию b-кетоацил-КоА. Заключительный этап катализирует тиолаза, расщепляющая тиосвязь с участием HS-Коэнзима-А, что приводит к образованию ацетил-КоА и ацил-КоА с укороченной на два углеродных атома цепью.
Ацетильный остаток ацетил-КоА окисляется в цикле Кребса, укороченная жирная кислота вовлекается в повторный цикл b-окисления. В конечном счете, она распадается на такое количество остатков ацетила, которое в два раза меньше числа атомов углерода в ней.
При окислении жирных кислот с нечетным количеством углеродных атомов на последнем цикле b-окисления образуется ацетил-КоА и активная форма пропионовой кислоты (пропионил-КоА). Пропионил-КоА карбоксилируется до метилмалонил-КоА и изомеризуется в сукцинил-КоА. Последний поступает в ЦТК.
Окисление жирных кислот

Синтез триглицеридов
Триглицериды синтезируются в стенке кишечника, в печени и жировой ткани (в адипоцитах).
Синтез триглицеридов в стенке кишечника может происходить из моноглицерида (из 2-моноацилглицерола) и двух молекул активных жирных кислот (остатки жирных кислот в комплексе с ацилпереносящим энзимом – S-КоА), или из глицерина и трех молекул активных жирных кислот с участием АТФ, что более характерно для процессов в печени и жировой ткани.
Синтез жира из глицерина и жирных кислот в печени и жировой ткани происходит по следующему пути. Глицерин фосфорилируется с использованием АТФ до глицерофосфата (фермент глицеролкиназа), затем под действием фермента глицеролфосфатацилтрансферазы взаимодействует с двумя молекулами ацилкоэнзима-А (например, с пальмитил-КоА). Образуется фосфатидная кислота (3-фосфо-1,2-
диацилглицерол). При взаимодействии последней с ацилкоэнзимом- А образуется триглицерид, свободный HS-КоА и остаток ортофосфорной кислоты.
Печень – основной орган, где идет синтез жирных кислот из продуктов гликолиза. Основной путь синтеза триглицелидов в печени из жирных кислот и глицерофосфата. Глицерофосфат, в свою очередь, в печень поступает из гидролиза жиров, а так же при восстановлении диоксиацетонфосфата (из гликолиза) при помощи восстановленного НАДФ (НАДФ Н+Н).

Окисление глицерина
Глицерин сначала фосфорилируется с участием АТФ до глицерофосфата (3- фосфоглицерол).
Затем под действием НАД-зависимой
глицерофосфатдегидрогеназы окисляется до 3-фосфоглицеринового альдегида.
Фосфоглицериновый альдегид далее может окисляться до пировиноградной и молочной кислоты.

Синтез жирных кислот
Синтез жирных кислот протекает в цитозоле из Ацетил-КоА, образовавшегося в митохондриях при гликолизе. Для использования ацетилкоэнзима-А в процессах, протекающих в цитоплазме клетки, протекает ряд реакций для переноса Ацетил-КоА через митохондриальную мембрану.
В митохондриях ацетил-КоА взаимодействует со Щавелевоуксусной кислотой (ЩУК), образуется лимонная кислота. В цитоплазме протекает обратный процесс. Таким образом, в цитоплазме образуется Ацетил-КоА.
Для синтеза жирных кислот протекает еще ряд последовательных реакций, образуется малонил-КоА. Ацетильная и малонильная группы переносятся на АПБ при участии ацетил-и малонил- трансацилаз. АПБ — ацилпереносящий белок. Далее к Ацетил-АПБ прибавляется Малонил-АПБ с образованием Ацетоацетил-АПБ. Наступает цикл реакций, противоположных бета-окислению жирных кислот, но вместо КоА носителем является АПБ, а вместо НАД и ФАД в процессах гидрирования участвует НАДФН2.
Бутирил-АПБ вступает в новый цикл (взаимодействует с малонил- АПБ), в результате которого углеродная цепь удлиняется на 2 атома. Циклы повторяются, пока цепь включит 16 атомов углерода (пальмитиновая кислота) или большего четного числа.

Метаболизм кетоновых тел
Кетоновые тела, являясь продуктами жирового обмена, выполняют в организме функцию энергетического субстрата, т.е. источника энергии. К кетоновым телам относят b-гидроксимасляную кислоту, ацетоуксусную кислоту и, как конечный (тупиковый) продукт – ацетон (диметилкетон). Это небольшие водорастворимые молекулы. Общая концентрация кетоновых тел в крови млекопитающих составляет около 5 мг% (у жвачных животных до 10 мг%). Синтезируются кетоновые тела в печени (в гепатоцитах) из ацетил-КоА. У жвачных животных отмечают также синтез кетоновых тел в слизистой оболочке преджелудков.
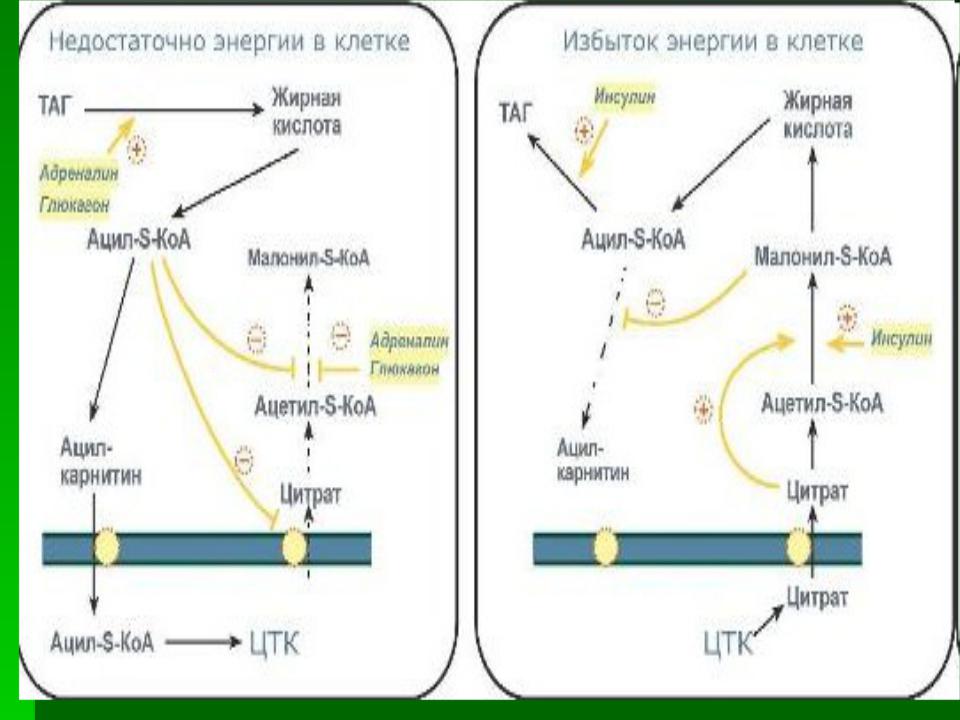
При выраженном углеводном голодании (белковый и белково-жировой рацион с недостатком углеводов), при нарушениях обмена углеводов (сахарный диабет) в результате клеточного голодания усиливается распад и использование жиров, что приводит к накоплению ацетил-КоА. Излишек
молекул ацетил-КоА вовлекается в синтез кетоновых тел. Резкое увеличение концентрации кетоновых тел в крови приводит к развитию кетозов и кетоацидозов.
Для синтеза кетоновых тел используется три молекулы ацетил-КоА. При конденсации двух молекул ац.-КоА образуется ацетоацетил-КоА; затем добавление третьей молекулы приводит к образованию b-гидрокси-b-метил- глютарил-КоА. При этих реакциях освобождается две молекулы HS-КоА. Полученная молекула b-гидрокси-b-метил-глютарил-КоА под действием лиазы распадается на ацетоуксусную кислоту и ацетил-КоА. Все указанные реакции протекают в матриксе митохондрий гепатоцитов.
В цитоплазме клеток ацетоуксусная кислота при участии НАД-зависимой дегидрогеназы (НАДФ*Н2) может восстанавливаться до b-гидроксимасляной кислоты или спонтанно декарбоксилироваться с образованием ацетона (диметилкетона).