

Шпоры / бббхххх
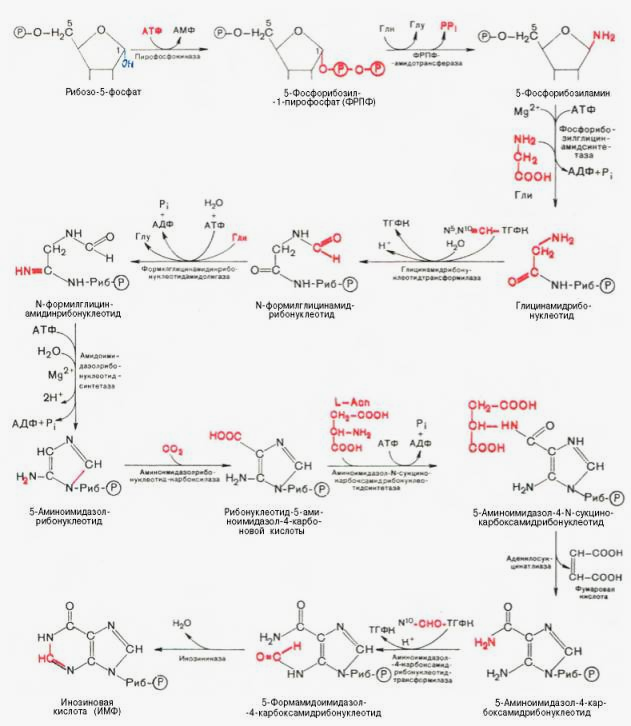
.docПредставлене о синтезе пуриновых нуклеотидов.
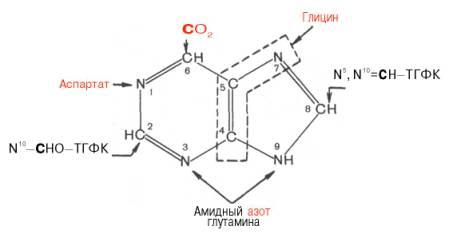
синтез инозиновой кислоты начинается с D-рибозо-5-фосфата, который является продуктом пентозофосфатного цикла и на который переносится пирофосфатная группа АТФ. Образовавшийся 5-фосфорибозил-1-пирофос-фат (ФРПФ) взаимодействует с глутамином, являющимся донором NH2-группы, в результате чего образуется β-5-фосфорибозил-амин, причем в процессе реакции наряду с освобождением пирофосфата и свободной глутаминовой кислоты происходит изменение его конфигурации (из α- в β-). Таким образом, данная стадия становится ключевой реакцией в синтезе пуринов
Представление о распаде и синтезе пиримидиновых нуклеотидов
 I
стадия синтеза УМФ включает катализируемое
цито-плазматической карбамоилфосфатсинтетазой
образование карбамоилфосфата из
глутамина
I
стадия синтеза УМФ включает катализируемое
цито-плазматической карбамоилфосфатсинтетазой
образование карбамоилфосфата из
глутамина
На II стадии карбамоилфосфат реагирует с аспартатом, в результате чего образуется N-карбамоиласпарагиновая кислота, она подвергается циклизации (под действием дигидрооротазы) с отщеплением молекулы воды, при этом образуется дигидрооротовая кислота, которая, подвергаясь дегидрированию, превращается в оротовую кислоту. В этой реакции участвует специфический НАД-содержащий фермент дигидро-оротатдегидрогеназа. Оротовая кислота обратимо реагирует с ФРПФ, являющимся донатором рибозо-фосфата, с образованием оротидин-5'-фос-фата (ОМФ). Декарбоксилирование последнего приводит к образованию первого пиримидинового нуклеотида – уридин-5-фосфата (УМФ).
Превращение УМФ в УДФ и УТФ осуществляется, как и пуриновых нуклеотидов, путем фосфотрансферазных реакций:
УМФ + АТФ <=> УДФ + АДФ ;
УДФ + АТФ <=> УТФ + АДФ.
Н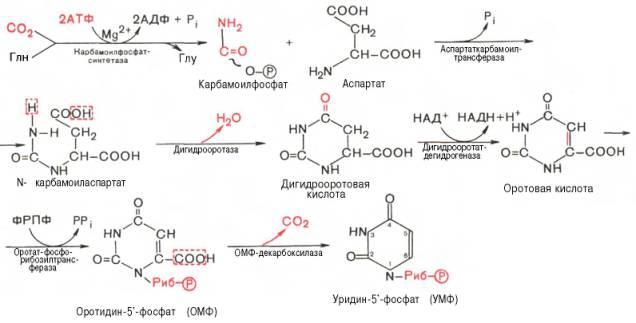 ачальные
этапы реакции распада пиримидиновых
нуклеотидов катализируются специфическими
ферментами. Конечными продуктами реакции
являются СО2, NH3, мочевина, β-аланин и
β-аминоизомасляная кислота. гидролитический
путь распада пиримидинов является
главным путем образования β-аланина,
который может служить источником для
синтеза ансерина и карнозина а также
для образования КоА
ачальные
этапы реакции распада пиримидиновых
нуклеотидов катализируются специфическими
ферментами. Конечными продуктами реакции
являются СО2, NH3, мочевина, β-аланин и
β-аминоизомасляная кислота. гидролитический
путь распада пиримидинов является
главным путем образования β-аланина,
который может служить источником для
синтеза ансерина и карнозина а также
для образования КоА
Регуляция биосинтеза пуриновых и пиримидиновых нуклеотидов.
Скорость синтеза нуклеотидов должна соответствовать потребностям клетки, в связи с чем она должна эффективным образом регулироваться. В работе механизмом регуляции синтеза пуриновых и пиримидиновых нуклеотидов много общего: решающую роль в регуляции играет ретроингибирование - снижение скорости синтеза нуклеотидов при достижении их достаточной концентрации в клетках за счет аллостерического ингибирования ключевых ферментов соответствующих метаболических путей.
Основными регуляторными ферментами метаболического пути синтеза пиримидиновых нуклеотидов являются карбамоилфосфатсинтетаза ( Е1 ) и аспартаттранскарбамоилаза ( Е2 ). Активность первого фермента ( Е1 ) ингибируется по аллостерическому механизму высокими концентрациями УТФ в клетке, а активность второго фермента ( Е2 ) - высокими концентрациями ГТФ. Активность карбамоифосфатсинтетазы, кроме того, активируется высокими концентрациями ФРПФ. С другой стороны, синтез ФРПФ тормозится высокими концентрациями дТДФ за счет аллостерического ингибирования ФРПФ-синтетазы ( Е3).
Нарушения обмена нуклеотидов.
Наиболее известным заболеванием, тесно связанным с нарушением обмена пуриновых нуклеотидов, является подагра. У больных с этой патологией наблюдается повышенное содержание мочевой кислоты в крови и тканях, а также избыточное количество уратов в моче. В норме концентрация мочевой кислоты в крови и других биологических жидкостях достаточно близка к насыщающей. Поэтому повышение ее содержания в биологических жидкостях приводит к появлению в них кристаллов мочевой кислоты. Если кристаллы появляются в суставной жидкости, развивается подагрические артриты. Выпадение кристаллов мочевой кислоты непосредственно в ткани вызывает асептическое воспаление с последующим инкапсулированием образовавшихся кристаллов и формированием подагрических узелков. Наиболее тяжелым проявлением этого заболевания является подагрическая нефропатия с нарушением функции почек.
При лечении подагры стремятся уменьшить в рационе количество продуктов, содержащих нуклеиновые кислоты или соединения группы пурина. Хороший эффект дает использование лекарственного препарата - аллопуринола. Аллопуринол в клетках под действием фермента ксантиноксидазы окисляется до аллоксантина, а аллоксантин является мощным конкурентным ингибитором ксантиноксидазы. Образование ксантина и мочевой кислоты в клетках резко снижается, а из организма в качестве конечного продукта обмена пуринов начинает выделяться гипоксантин, растворимость которого в биологических жидкостях в несколько раз выше, чем растворимость мочевой кислоты.
Биосинтез дезокстирибонуклеотидов
Синтез дезоксирибонуклеотидов происходит из рибонуклеозиддифосфатов путем восстановления рибозы в составе нуклеотида в дезоксирибозу. В этом процессе участвуют три белка:
рибонуклеозидредуктаза — фермент, катализирующий восстановление рибозы в составе нуклеотида
тиоредоксин — донор водорода для восстановления кислорода рибозы в положении С—2 до молекулы воды
тиоредоксинредуктаза — фермент, восстанавливающий окисленный тиоредоксин за счет водорода NADFH
Дезоксинуклеозиддифосфаты затем превращаются в дезоксинуклеозидтрифосфаты и служат субстратами для синтеза DNA. Так синтезируются dATP, dGTP, dCTP. В результате метилирования dTTP синтезируется из dUMP. Донором одноуглеродной группы — СН3 служит метилен—ТГФ. Реакция катализируется тимидилатсинтазой
В ходе превращения образуется дигидрофолат, который восстанавливается снова до тетрагидрофолата под действием фолатредуктазы и водорода NADFH. На ингибировании тимидилатсинтетазы и фолатредуктазы основано действие противоопухолевых препаратов. Эти препараты тормозят образование тимидина из dTTP, необходимых для синтеза DNA, а, следовательно, репликацию и деление клеток.