

1. Работа калий-натриевого насоса мембраны.
Новая гипотеза механизма работы Na,K-АТФазы рассматривается здесь: Механизм натрий-калиевого насоса
Работа калий-натриевого насоса, в свою очередь, имеет 2 следствия:
1.1. Непосредственное электрогенное (порождающее электрические явления) действие ионного насоса-обменника. Это создание небольшой электроотрицательности внутри клетки (-10 мВ).
Виноват в этом неравный обмен натрия на калий. Натрия выбрасывается из клетки больше, чем поступает в обмен калия. А вместе с натрием удаляется и больше "плюсиков" (положительных зарядов), чем возвращается вместе с калием. Возникает небольшой дефицит положительных зарядов. Мембрана изнутри заряжается отрицательно (примерно -10 мВ).
1.2. Создание предпосылок для возникновения большой электроотрицательности.
Эти предпосылки - неравная концентрация ионов калия внутри и снаружи клетки. Лишний калий готов выходить из клетки и выносить из неё положительные заряды. Об этом мы скажем сейчас ниже.
2. Утечка ионов калия из клетки.
Из зоны повышенной концентрации внутри клетки ионы калия выходят в зону пониженной концентрации наружу, вынося заодно положительные электрические заряды. Возникает сильный дефицит положительных зарядов внутри клетки. В итоге мембрана дополнительно заряжается изнутри отрицательно (до -70 мВ).
Финал
Итак:
Калий-натриевый насос создает предпосылки для возникновения потенциала покоя. Это - разность в концентрации ионов между внутренней и наружной средой клетки. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка клетки выравнять концентрацию ионов по калию приводит к потере калия, потере положительных зарядов и порождает электроотрицательность внутри клетки. Эта электроотрицательность составляет большую часть потенциала покоя. Меньшую его часть составляет непосредственная электрогенность ионного насоса, т.е. преобладающие потери натрия при его обмене на калий.
Мембранный потенциал покоя
Мембранным потенциалом покоя (МПП) или потенциалом покоя (ПП) называют разность потенциалов покоящейся клетки между внутренней и наружной сторонами мембраны. Внутренняя сторона мембраны клетки заряжена отрицательно по отношению к наружной. Принимая потенциал наружного раствора за нуль, МПП записывают со знаком «минус». Величина МПП зависит от вида ткани и варьирует от -9 до -100 мв. Следовательно, в состоянии покоя клеточная мембрана поляризована. Уменьшение величины МПП называютдеполяризацией, увеличение — гиперполяризацией, восстановление исходного значения МПП —реполяризацией мембраны.
Основные положения мембранной теории происхождения МПП сводятся к следующему. В состоянии покоя клеточная мембрана хорошо проницаема для ионов К+ (в ряде клеток и для СГ), менее проницаема для Na+и практически непроницаема для внутриклеточных белков и других органических ионов. Ионы К+диффундируют из клетки по концентрационному градиенту, а непроникающие анионы остаются в цитоплазме, обеспечивая появление разности потенциалов через мембрану.
Возникающая разность потенциалов препятствует выходу К+ из клетки и при некотором ее значении наступает равновесие между выходом К+ по концентрационному градиенту и входом этих катионов по возникшему электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, называется равновесным потенциалом. Его величина может быть рассчитана из уравнения Нернста:
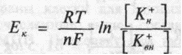
где Ек — равновесный потенциал для К+; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; п — валентность К+ (+1), [Кн+] — [К+вн] — наружная и внутренняя концентрации К+-
Если
перейти от натуральных логарифмов к
десятичным и подставить в уравнение
числовые значения констант, то уравнение
примет вид:
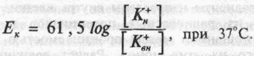
В спинальных нейронах (табл. 1.1) Ек = -90 мв. Величина МПП, измеренная с помощью микроэлектродов заметно ниже — 70 мв.
Таблица 1.1. Концентрация некоторых ионов внутри и снаружи спинальных мотонейронов млекопитающих
|
Ион |
Концентрация |
(ммоль/л Н2О) |
Разновесный потенциал (мв) |
|
внутри клетки |
снаружи клетки | ||
|
Na+ |
15,0 |
150,0 |
+60 |
|
К+ |
150,0 |
5,5 |
-90 |
|
Сl- |
9,0 |
125,0 |
-70 |
|
|
Мембранный потенциал покоя = -70 мв | ||
Если мембранный потенциал клетки имеет калиевую природу, то, в соответствии с уравнением Нернста, его величина должна линейно снижаться с уменьшением концентрационного градиента этих ионов, например, при повышении концентрации К+ во внеклеточной жидкости. Однако линейная зависимость величины МПП (Мембранный потенциал покоя) от градиента концентрации К+ существует только при концентрации К+во внеклеточной жидкости выше 20 мМ. При меньших концентрациях К+ снаружи клетки кривая зависимости Ем от логарифма отношения концентрации калия снаружи и внутри клетки отличается от теоретической. Объяснить установленные отклонения экспериментальной зависимости величины МПП и градиента концентрации К+ теоретически рассчитанной по уравнению Нернста можно, допустив, что МПП возбудимых клеток определяется не только калиевым, но и натриевым, и хлорным равновесным потенциалами. Рассуждая аналогично с предыдущим, можно записать:
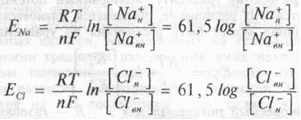
Величины натриевого и хлорного равновесных потенциалов для спинальных нейронов (табл. 1.1) равны соответственно +60 и -70 мв. Значение ЕCl равно величине МПП. Это свидетельствует о пассивном распределении ионов хлора через мембрану в соответстии с химическим и электрическим градиентами. Для ионов натрия химический и электрический градиенты направлены внутрь клетки.
Вклад каждого из равновесных потенциалов в величину МПП определяется соотношением между проницаемостью клеточной мембраны для каждого из этих ионов. Расчет величины мембранного потенциала производится с помощью уравнения Гольдмана:

Еm — мембранный потенциал; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; РK , PNa и РCl — константы проницаемости мембраны для К+ Na+ и Сl, соответственно; [К+н], [K+вн, [Na+н [Na+вн], [Сl-н] и[Сl-вн ]- концентрации K+, Na+ и Сl снаружи (н) и внутри (вн) клетки.
Подставляя в это уравнение полученные в экспериментальных исследованиях концентрации ионов и величину МПП, можно показать, что для гигантского аксона кальмара должно быть следующее соотношение констант проницаемости Рк : PNa : РС1 = I : 0,04 : 0,45. Очевидно, что, поскольку мембрана проницаема для ионов натрия (РNa =/0) и равновесный потенциал для этих ионов имеет знак «плюс», то вход последних внутрь клетки по химическому и электрическому градиентам будет уменьшать электроотрицательность цитоплазмы, т.е. увеличивать МПП (Мембранный потенциал покоя).
При повышении концентрации ионов калия в наружном растворе выше 15 мМ МПП увеличивается и соотношение констант проницаемости меняется в сторону более значительного превышения» Рк над PNa и РС1. Рк: PNa : РС1 = 1 : 0.025 : 0,4. В таких условиях МПП определяется почти исключительно градиентом ионов калия, поэтому экспериментальная и теоретическая зависимости величины МПП от логарифма отношения концентраций калия снаружи и внутри клетки начинают совпадать.
Таким образом, наличие стационарной разности потенциалов между цитоплазмой и наружной средой в покоящейся клетке обусловлено существующими концентрационными градиентами для К+, Na+ и Сl и различной проницаемостью мембраны для этих ионов. Основную роль в генерации МПП играет диффузия ионов калия из клетки в наружный наствор. Наряду с этим, МПП определяется также натриевым и хлорным равновесными потенциалами и вклад каждого из них определяется отношениями между проницаемостями плазматической мембраны клетки для данных ионов.
Все факторы, перечисленные выше, составляют так называемую ионную компоненту МПП (Мембранный потенциал покоя). Поскольку, ни калиевый, ни натриевый равновесные потенциалы не равны МПП. клетка должна поглощать Na+ и терять К+. Постоянство концентраций этих ионов в клетке поддерживается за счет работы Na+ К+-АТФазы.
Однако роль этого ионного насоса не ограничивается поддержанием градиентов натрия и калия. Известно, что натриевый насос электрогенен и при его функционировании возникает чистый поток положительных зарядов из клетки во внеклеточную жидкость, обуславливающий увеличение электроотрицательности цитоплазмы по отношению к среде. Электрогенность натриевого насоса была выявлена в опытах на гигантских нейронах моллюска. Электрофорети-ческая инъекция ионов Na+ в тело одиночного нейрона вызывала гиперполяризацию мембраны, во время которой МПП был значительно ниже величины калиевого равновесного потенциала. Указанная гиперполяризация ослаблялась при снижении температур раствора, в котором находилась клетка, и подавлялась специфическим ингибитором Na+, К+-АТФазы уабаином.
Из сказанного следует, что МПП может быть разделен на две компоненты — «ионную» и «метаболическую». Первая компонента зависит от концентрационных градиентов ионов и мембранных проницаемостей для них. Вторая, «метаболическая», обусловлена активным транспортом натрия и калия и оказывает двоякое влияние на МПП. С одной стороны, натриевый насос поддерживает концентрационные градиенты между цитоплазмой и внешней средой. С другой, будучи электрогенным, натриевый насос оказывает прямое влияние на МПП. Вклад его в величину МПП зависит от плотности «насосного» тока (ток на единицу плошади поверхности мембраны клетки) и сопротивления мембраны.