

Нейрофизиология / polyarizatsionnaya-teoriya-motivatsiy-emotsiy-i-vnimaniya
.pdfБЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45)
У Д К 3 1 6 . 6 2 8
С.Э. Мурик
ПОЛЯРИЗАЦИОННАЯ ТЕОРИЯ МОТИВАЦИЙ, ЭМОЦИЙ И ВНИМАНИЯ
Иркутский государственный университет (Иркутск)
Предлагается новая теория мотиваций, эмоций и внимания, рассматривающая их как функции сен сорных систем и связывающая нейрофизиологический механизм указанных психических феноменов с изменением метаболического и функционального состояния перцептивных нейронов, отражаю щегося на степени стационарной поляризации мембраны клеток.
Ключевые слова: мотивации, эмоции, внимание, нейрофизиологический механизм
POLARIZATION THEORY OF MOTIVATION, EMOTIONS AND ATTENTION
S.E. Murik
Irkutsk State University, Irkutsk
The new theory of motivation, emotions and attention is suggested, which considers them as functions of sensory systems and connects neurophysiological mechanism of mentioned mental phenomena with the change of metabolic and functional condition of perceptive neurons, reflecting on the degree of permanent polarization of cells membrane.
Key words: motiv ations, emotions, attention, neurophysiological mechanism
Одной из фундаментальных проблем совре менного естествознания является проблема физи ологической основы психической деятельности человека и животных. Состояние решения данно го вопроса в настоящее время вряд ли можно при знать удовлетворительным. За последние 100 лет на фоне очевидных успехов таких биологических наук как генетика, молекулярная биология, био физика и других, физиологией психики по сути не решено ни одной стоящей перед ней задачи. Се годня мы, как и более столетия назад, не знаем реального механизма ни одного психического фе номена. Все наши знания заключаются в огромном количестве накопленных экспериментальных дан ных, с трудом укладываемых в схемы, которые с большой осторожностью можно назвать реальным устройством и механизмами работы мозга.
«Мысль есть первые две трети психического рефлекса», – сказал, в свое время, И.М. Сеченов [38]. Ничего больше к тому, что сказал И.М. Сече нов о нейрональном механизме мышления, мы и сегодня добавить не можем.
Одни из первых естественнонаучных идей о механизмах эмоций и мотиваций принадлежат W. James [50] и W.B. Cannon [47]. Суть их заклю чалась в том, что и то, и другое явление – резуль тат перцепции мозгом раздражителей. В настоя щее время, наука на 100 % уверовала в то, что дан ные феномены, на самом деле, рождаются в не рвной системе, но каков их нейрональный меха низм мы определенно так и не знаем.
Механизм мотиваций, в настоящее время, рас сматривается в контексте существования специфи ческих и неспецифических мотивациогенных цен тров и «центрального мотивационного возбужде ния» [39, 48, 56, 61, 63]. Несмотря на длительную
историю проблемы физиологии мотиваций сегод ня все еще остаются недостаточно ясными приро да мотивационного возбуждения и нейрофизиоло гические механизмы мотивированных состояний.
Неудовлетворительным является также со стояние проблемы физиологической основы эмо ций. Как и двадцать лет назад, сегодня актуаль ным остается высказывание немецких физиоло гов Р. Шмидта и Г. Тевса [49] о том, что в настоя щее время не существует единой общепринятой научной теории эмоций, а также точных данных о том, в каких центрах и каким образом эти эмо ции возникают и каков их нервный субстрат.
Особенностью современного подхода к про блеме мотиваций и эмоций является рассмотрение их механизма вне сенсорных систем и, соответ ственно, вне перцептивных процессов [40, 52, 53, 62]. Данный подход определяется доминировани ем в физиологии психической деятельности ки бернетических идей [65]. Суть этого подхода зак лючается в выделении в мозге самостоятельных структурно функциональных элементов, их упо рядочение и соподчинение по механистическим принципам. Исследование мозга под углом данно го мировоззрения предполагает вычленение в моз ге систем управления и контроля и внедрение их в механизмы работы мозга.
Современная физиология мотиваций и эмоций не только исключает их механизмы из акта пер цепции, но и в самих этих механизмах выделяет отдельные феномены (подфункции) с самостоя тельными мозговыми субстратами [15, 39, 40]. В конечном итоге, мозг становится бесконечно сложной механической «матрешкой», в которой та или иная система состоит из нескольких элемен тов, а те, в свою очередь, из собственных элемен
Экспериментальные исследования в медицине и биологии |
167 |

БЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45)
тов и т.д. Установить характер взаимосвязей и от ношений в такой системе между множеством эле ментов разного уровня становится бесконечно сложной задачей. Поэтому в познании механизмов психической деятельности, к настоящему време ни, практически прекратилось движение вперед.
Выход, на наш взгляд, лежит в отказе от сис темного [4] и кибернетического [65] принципов в решении проблемы структурно функциональной организации мозга и рассмотрении нейрофизио логического механизма мотиваций, эмоций и вни мания в связи с изменением функционального со стояния (ФС) афферентных (перцептивных) ней ронов [20–30, 32–35].
В настоящее время термин ФС широко исполь зуется в физиологии при изучении различных уровней организма человека и животных, однако
внего при этом может вкладываться совершенно разный смысл. В нейрофизиологии, говоря о ФС нейронов, подразумевают такие явления как воз буждение, торможение и покой [7, 13, 12, 19].
Функциональное же состояние в буквальном смысле этих слов означает состояние функции или состояние выполнения функции. Отвечая на воп рос о ФС, необходимо описать, в каком состоянии находится выполнение той или иной функции: в хорошем или плохом, т.е. дать качественную ха рактеристику функции [17, 18].
Исходя из представления о ФС как качествен ной характеристики деятельности, функциональ ное состояние нейрона может быть определено как показатель способности клетки выполнять свой ственные ей функции на данный момент времени.
Для нейрона основной функцией, которую он выполняет, является генерация потенциала действия (нервного импульса). Мерой эффек тивности выполнения этой функции будет ла бильность (или функциональная подвижность), т.е. количество импульсов, которое клетка мо жет генерировать в единицу времени [7]. Рас смотрение показателя лабильности в качестве эффективностной меры деятельности нейронов
вфизиологии в настоящее время практически не встречается, однако именно лабильность, по сути, отражает ФС нейрона.
Анализ литературных данных показывает, что эффективность деятельности нейрона (функци ональная способность или лабильность) меняет ся при изменении уровня мембранного потенци ала (МП). Деполяризация сопровождается сниже нием, а гиперполяризация повышением лабиль ности возбудимого образования [42], соответ ственно этому меняется и ФС нейрона: при гиперполяризации – улучшается, при деполяри зации – ухудшается.
Устойчивая деполяризация МП является не благоприятным не только с функциональной, но и с метаболической точки зрения, поскольку появ ление устойчивой (стационарной) деполяризации нейрональной мембраны в процессе нервной дея тельности может произойти только в том случае, если механизмы, восстанавливающие ионный го
меостаз после генерации нервных импульсов, не справляются со своей задачей. Давно показано [44], что состояние макроэргических систем акти вированных нервных клеток быстро истощается и становится похожим на состояние энергетичес кого голода. Иначе говоря, развитие устойчивой деполяризации потенциала покоя нейрона в про цессе импульсной активности отражает появление дефицита макроэргических соединений. Показа но также, что устойчивая деполяризация МП за пускает целый каскад патогенетических процес сов, приводящих в конечном итоге клетки к гибе ли [8, 11, 51, 55, 64].
Таким образом, в настоящее время, нельзя не признать, что развитие устойчивой деполяризации потенциала покоя клетки в промежутках между ПД не является индифферентным с точки зрения качества метаболизма состоянием, а свидетель ствует о преобладании процессов катаболизма и отставании анаболических процессов от жизнен ных потребностей клетки, т.е. об ухудшении, в целом, в ней жизненных явлений.
Как говорилось выше, лабильность нейрона увеличивается при гиперполяризации МП [42]. Анализ литературных данных [2, 41, 45] также по казывает, что активации нейронов в связи с пере работкой информации из внешней и внутренней среды организма, как правило, предшествует именно гиперполяризационное отклонение МП, а генерация ПД выглядит как постгиперполяризаци онная отдача. Следовательно, гиперполяризация может рассматриваться, как отражение рабочего ФС нервных клеток. Поскольку гиперполяризаци онное отклонение МП является обязательной пер вичной реакцией нервных клеток на любые воз действия [6, 7], то формирующееся при этом со стояние, по всей видимости, отражает мобилиза цию адаптивных внутриклеточных механизмов. О том, что во время гиперполяризации мобилизуют ся адаптивные резервы клеток, говорят экспери ментальные данные, свидетельствующие об увели чении в этот период их резистентности [37]. Из вестно также, что гиперполяризующие препара ты обладают протекторным действием в неблагоп риятных для жизни нейронов условиях [16].
В настоящее время можно с уверенностью ут верждать, что ФС нейрона (лабильность) зависит от способности структуры восстанавливать свои энергетические и пластические запасы, необходи мые для осуществления полноценной реакции. Иначе говоря, ФС как характеристика эффектив ности выполнения нейронами их функций тесно связано с метаболическими процессами в клетках. Выполнение нейронами функций влияет на их метаболическое состояние, сдвиги которого в свою очередь определяют функциональные воз можности клеток. Поэтому сдвиги ФС всегда вза имосвязаны со сдвигами метаболического состо яния, а показатели ФС характеризуют и метабо лическое состояние возбудимого образования. Следовательно, изменение лабильности может рассматриваться в качестве индикатора сдвигов
168 |
Экспериментальные исследования в медицине и биологии |

БЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45)
функционального и метаболического состояния возбудимой ткани. Снижение лабильности свиде тельствует о плохом, а увеличение – о хорошем функциональном и метаболическом состоянии данного нервного образования.
Таким образом, анализ литературных данных свидетельствует о том, что интегральным показа телем метаболического и ФС нейронов мозга яв ляется уровень МП. Совокупность биохимических и электрофизиологических данных указывает так же на то, что устойчивая деполяризация МП кле ток отражает развитие в них неблагоприятного функционального и метаболического состояния, и поэтому должна быть нежелательна как для от дельных клеток, так и организма в целом. Разви тие неблагоприятного ФС в популяциях нейронов, в этом случае, может лежать в основе выявления нервной системой раздражителей и факторов, имеющих отрицательное биологическое значение для организма и организации на этой основе адап тивного поведения. Иначе говоря, биологическая значимость раздражителя может быть оценена по ФС воспринимающих его нейронов.
Развитие неблагоприятного ФС, типа устойчи вой (стационарной) деполяризации может являть ся нейрофизиологической основой мотивирован ных состояний, и субъективного переживания в виде негативных эмоций. Тогда устранение в орга низме данного состояния, т.е. реполяризация и гиперполяризация нейронов посредством гумо ральных или поведенческих реакций, будет награ дой и механизмом положительных эмоций.
Исследование поляризационных процессов в нервной системе возможно посредством регист рации так называемого постоянного потенциала (steady potential или direct current potential [46, 54].
При этом негативный сдвиг постоянного потенци ала отражает деполяризацию МП клеток нервной ткани, тогда как позитивный обратный процесс – реполяризацию или гиперполяризацию. Комплек сная регистрация уровня постоянного потенциа ла (УПП) и ЭЭГ может быть хорошим методом оценки функционального и метаболического со стояния нервной ткани головного мозга [31, 59].
Как показали проведенные нами эксперимен ты на крысах [30, 54, 57] при мотивированных со стояниях и сопровождающих их негативных эмо циях в нервной ткани головного мозга развива ются деполяризационные процессы, сходные с теми, которые наблюдаются при неглубокой ише мии [36].
Как видно на рис. 1 относительно слабая ише мия мозга, связанная с частичным ограничением мозгового кровотока («Ишемия 1»), сопровожда лась увеличением мощности ритмов ЭЭГ и неболь шой негативизацией постоянного потенциала. В среднем снижение потенциала по всей выборке (n = 24) составило 1222,51 ± 290,1 мкВ. Еще боль шее ограничение мозгового кровотока посред ством дополнительного перекрытия средней моз говой артерии («Ишемия 2») привело на фоне зна чительной негативизации постоянного потенциа ла к депрессии ЭЭГ.
На рис. 2 показано изменение тех же самых показателей после введения крысам в брюшную полость наркотического вещества.
Видно, что сразу после инъекции препарата наблюдалось негативное отклонение УПП и уве личение мощности ЭЭГ. Негативизация потенци ала составила в среднем 638,8 ± 124,1 мкВ. Таким образом, негативное эмоциональное возбуждение, развивающееся, очевидно, у животных после фик
|
|
|
|
|
Теменная кора левого полушария |
|
|
|
|
|||||
|
|
|
Ишемия 1 |
|
|
|
Ишемия 2 |
|
|
|
Реперфузия |
|||
|
700 |
|
|
|
|
|
|
|
|
30000 |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
600 |
|
|
|
|
|
|
|
|
|
|
|
|
20000 |
|
500 |
|
|
|
|
|
|
|
|
|
|
|
|
10000 |
мкВ |
400 |
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
300 |
|
|
|
|
|
|
|
|
|
|
|
|
-10000 |
|
200 |
|
|
|
|
|
|
|
|
|
|
|
|
-20000 |
|
100 |
|
|
|
|
|
|
|
|
|
|
|
|
-30000 |
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
-40000 |
|
4 |
12 |
20 |
28 |
36 |
44 |
52 |
60 |
68 |
76 |
84 |
92 |
100 |
108 |
|
|
|
|
|
|
|
|
мин |
|
|
|
|
|
|
|
|
|
Дельта-1 |
Дельта-2 |
|
Тета |
Альфа |
Бета |
|
УПП |
|
|||
Рис. 1. Изменение УПП и ЭЭГ в теменной коре левого полушария у крысы при моделировании ишемии мозга разными |
||||||||||||||
|
способами [36]. Обозначения: «Ишемия 1» – перевязывание общих сонных артерий; «Ишемия 2» – введение |
|||||||||||||
|
окклюдера в среднюю мозговую артерию левого полушария (СМА); «Реперфузия» – извлечение окклюдера из |
|||||||||||||
|
СМА. На левой шкале оси Y отложена амплитуда ритмов ЭЭГ (mкВ), на правой – УПП (mкВ). Значения УПП в |
|||||||||||||
|
период, предшествовавший «Ишемии01», принят за ноль. |
|
|
|
|
|
|
|||||||
Экспериментальные исследования в медицине и биологии |
169 |
БЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45)
сации их в руках, прокалывания иглой кожных покровов и введении препарата, сопровождалось негативизацией постоянного потенциала и увели чением мощности ЭЭГ, что в целом соответство вало электрофизиологическим изменениям, на блюдаемым во время «Ишемии 1» (рис. 1), и сви детельствует о сходстве явлений, развивающихся в обоих случаях на нейрональном уровне. В том и другом случае, по всей видимости, развивалось состояние деполяризационной экзальтации возбу димости нейронов и усиление метаболизма не рвной ткани по катэлектротоническому типу.
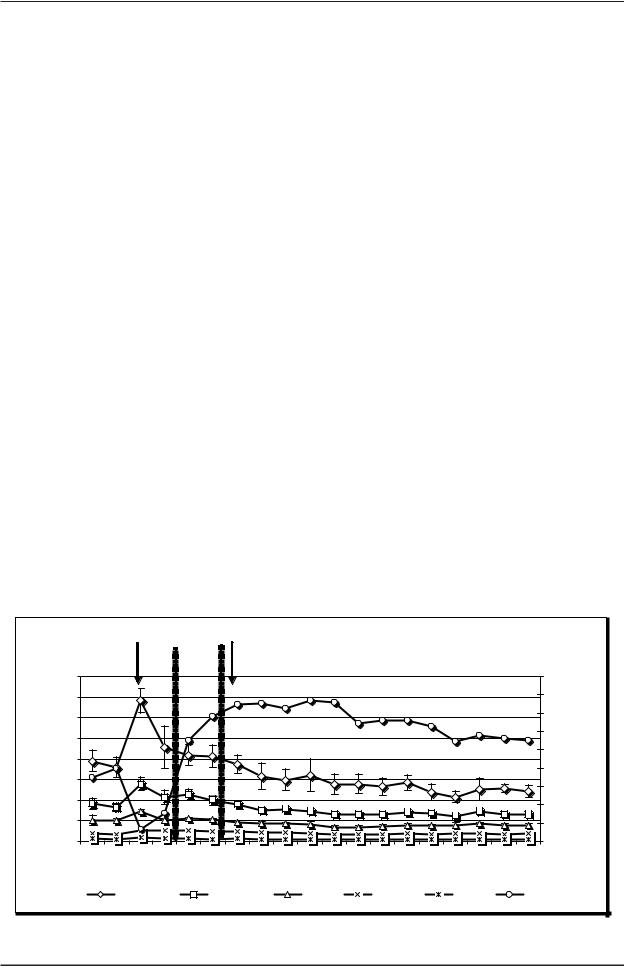
По мере всасывания в кровь этаминал натрия появилось позитивное отклонение УПП при сохра нении повышенной мощности ЭЭГ (на рис. 2 этот период обозначен как «Предсон»). Характер изме нения УПП (а именно его позитивизация) свиде тельствует о развитии при этом ре и гиперполя ризационных процессов. Увеличенная амплитуда ритмов указывает на отсутствии еще в это время гиперполяризационного торможения. Сопостав ление УПП и ЭЭГ характеристик позволяет рас сматривать развитие у нейронов в периоде «Пред сон» функционального состояния, соответствую щего анодной экзальтации, и усиленного метабо лизма нервной ткани. Наконец, наступление и раз витие наркотического сна сопровождалось еще более сильным позитивным сдвигом постоянного потенциала, сочетающегося с угнетением мощно сти ЭЭГ. Данное функциональное состояние от ражает, по всей видимости, углубление гиперпо ляризации клеток и наступление гиперполяриза ционного торможения со снижением метаболиз ма нервной ткани.
Видно, что в целом характер поляризационных процессов после ведения наркотика был противо положный тому, который наблюдался при ише
мии. Это указывает на то, что при этом развива лось другое метаболическое и ФС нейронов моз га: ишемия – модель формирования неблагопри ятного, а нембуталовый наркоз – хорошего фун кционального и метаболического состояния не рвной ткани.
Развитие деполяризационных процессов, от ражающих формирование неблагоприятного ме таболического и ФС, наблюдалось нами также у животных в состоянии голода. При формировании пищевой мотивации в структуре относимой к пи щевому центру (латеральное гипоталамическое ядро) деполяризация составляла 460 мкВ [57]. Про цесс потребления пищи и состояние пищевого насыщения сопровождались реполяризационны ми процессами в указанной структуре.
Как показали наши эксперименты (рис. 1 и 2) активация нейронов возможна, как на фоне депо ляризационных, так и гиперполяризационных процессов, т.е. как при плохом, так и хорошем их метаболическом и функциональном состоянии.
Генерация ПД по типу постгиперполяризаци онной отдачи является более эффективным с ме таболической точки зрения механизмом возбуж дения, чем какой либо другой и соответственно может рассматриваться как наиболее оптималь ный (рабочий) в нервной системе. Гиперполяри зационная волна, предшествующая возбужде нию, по всей видимости, мобилизует гомеостати ческие адаптивные механизмы. В этом состоянии клетка может генерировать большее число ПД и легче, чем в каком либо другом. Гиперполяриза ционное отклонение МП повышает функцио нальные возможности клетки и тем самым адап тивные возможности. Следовательно, так назы ваемое, гиперполяризационное торможение, по сути, и не торможение, а состояние повышенной
Введение |
|
|
Предсон |
|
Сон |
|
|
|
|
|
|
|
|
|
|
|
||
наркоза |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
400 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1200 |
350 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1000 |
300 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
800 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
600 |
|
250 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
400 |
|
мкВ 200 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
200 |
|
150 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
100 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-200 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
50 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-400 |
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-600 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
|
|
|
|
|
|
|
|
|
мин |
|
|
|
|
|
|
|
|
|
|
Дельта-1 |
|
Дельта-2 |
|
Тета |
|
Альфа |
|
Бета |
|
УПП |
|||||||
Рис. 2. Изменение уровня постоянного потенциала и амплитуды ритмов ЭЭГ после внутрибрюшинного введения нем0 бутала [59].
170 |
Экспериментальные исследования в медицине и биологии |
|
БЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45) |
|
резистентности и мобилизованности функцио |
его компонентов. В частности, нейрофизиологи |
|
нальных и метаболических ресурсов клетки. Если |
ческая основа внимания также, по всей видимо |
|
рассматривать его с качественной точки зрения, |
сти, тесно связана с функциональным состояни |
|
– это превосходное в функциональном отноше |
ем воспринимающих нейронов. Формирование |
|
нии ФС, благоприятное для выполнения свой |
в афферентных системах под действием раздра |
|
ственных для нервной клетки функций, в первую |
жителей энграмм, включающих нейроны с не |
|
очередь, генерации ПД. |
благоприятным функциональным состоянием, |
|
Таким образом, уравнивание, с функциональ |
будет, очевидно, «сигналом» для воспринимаю |
|
ной точки зрения, возбуждения и торможения, |
щей системы о необходимости организации |
|
рассмотрение их как равноценных рабочих актов |
«ориентировочного поведения», направленного |
|
не оправдано с биологических позиций. Состоя |
на устранение (редукцию) данного состояния. |
|
ние жизненных процессов при возбуждении (ка |
Иначе говоря, с позиций развиваемой концеп |
|
тэлектротоническом или постгиперполяризацион |
ции, внимание является разновидностью моти |
|
ном) и торможении (гиперполяризационном или |
вации. Таким образом, функциональное состоя |
|
деполяризационном) сильно различается. Поэто |
ние нейронов нервной модели стимула может |
|
му процессы реагирования на те или иные факто |
быть универсальным механизмом, лежащим в |
|
ры на уровне отдельных клеток (возбуждение и |
основе многих психофизиологических феноме |
|
торможение), скорее всего, необходимо рассмат |
нов, в том числе внимания. |
|
ривать как разные адаптивные акты с соответству |
До настоящего времени, мозговые механиз |
|
ющей характеристикой напряжения внутрикле |
мы указанных психических феноменов рассмат |
|
точных механизмов адаптации и качественными |
ривались вне анализаторных систем, что в конеч |
|
изменениями в них метаболизма. |
ном итоге и предопределяло отсутствие реальных |
|
Совокупность накопленных к настоящему |
подвижек в их понимании. Большинство извест |
|
времени физиологических данных неоспоримо |
ных теорий в соответствии с кибернетическим |
|
указывает на то, что устойчивая деполяризация |
(механистическим) подходом выделяло для них |
|
МП отражает развитие неблагоприятного мета |
самостоятельные мозговые аппараты. Все совре |
|
болического состояния, и поэтому не может яв |
менные популярные теории мозговой субстрат |
|
ляться желательным как для отдельных клеток, |
эмоций и мотиваций позициируют в так называ |
|
так и организма в целом. В этом случае, характер |
емой лимбической системе [1, 10, 39, 40]. Кибер |
|
поляризационных сдвигов должен обозначать |
нетическая идея о существовании в мозге само |
|
биологическую значимость раздражителей и из |
стоятельного нейронального аппарата эмоций и |
|
менений факторов среды. Развитие неблагопри |
мотиваций не находит достаточного фактическо |
|
ятного метаболического состояния будет свиде |
го обоснования [9]. Критический анализ струк |
|
тельствовать об отрицательных биологических |
турно функциональной организации лимбичес |
|
свойствах действующих агентов, тогда как хоро |
кой коры и тесно связанных с ней других облас |
|
шее метаболическое состояние, развивающееся |
тей головного мозга, включаемых в лимбическую |
|
под действием тех или иных факторов, – о поло |
систему [52, 53], не дает оснований для того, что |
|
жительных. |
бы считать их центральным нервным субстратом |
|
Развитие неблагоприятного метаболического |
субъективного переживания. Существующие |
|
и ФС в нейрональных системах, воспринимаю |
морфо функциональные данные [3, 5, 14] бес |
|
щих раздражители и изменения условий среды, |
спорно указывают лишь на то, что лимбическая |
|
не может не отразиться в субъективном мире |
система включает в себя нервные элементы вис |
|
индивида в появлении негативного эмоциональ |
церального анализатора и эфферентных систем, |
|
ного переживания. Поэтому совокупность имею |
регулирующих деятельность внутренних органов |
|
щихся литературных данных, а также получен |
и состояние внутренней среды организма. Ина |
|
ных нами, позволяет, с большой долей уверенно |
че говоря, лимбическая система если и имеет от |
|
сти, говорить, что изменение метаболического, а, |
ношение к эмоциям, то только к переживанию |
|
соответственно, и ФС перцептивных нейронов |
висцеральных стимулов или как нервный суб |
|
должно рассматриваться в качестве возможной |
страт вегетативных и отчасти соматических |
|
нейрофизиологической основы эмоций. Мобили |
врожденных эфферентных реакций, сопровож |
|
зация этими же нейронами системных механиз |
дающих эмоции. В таком случае, подход, посту |
|
мов адаптации в виде активации поведенческих |
лирующий наличие у эмоциональных (субъектив |
|
энграмм является основой и целенаправленного |
ных) переживаний самостоятельного нервного |
|
(мотивированного) поведения. Следовательно, |
центра, остается бездоказательным. |
|
нейрофизиологический механизм мотиваций и |
Разрабатываемая нами концепция ФС нейро |
|
эмоций, по всей видимости, слит с сенсорными |
нов и его роли в механизмах психической деятель |
|
процессами и заключается в изменении метабо |
ности, позволяет говорить о необходимости сме |
|
лического и ФС перцептивных нейронов. |
ны доминирующего в настоящее время механис |
|
Развиваемый нами подход к содержанию по |
тического подхода и к проблеме структурно функ |
|
нятия ФС на клеточном уровне дает возмож |
циональной организации мозга. Поскольку теку |
|
ность кардинально пересмотреть механизм це |
щее состояние нейрона, по сути, есть функция |
|
лостного поведенческого акта и составляющих |
производная от активации его адаптационных ме |
|
|
|
|
Экспериментальные исследования в медицине и биологии |
171 |
|

БЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45)
ханизмов, то вся нервная система и ее деятель |
тельно неглубокими сдвигами ФС в небольших |
|
ность может быть рассмотрена как совокупность |
объемах нервной ткани, приближающихся к не |
|
клеток адаптирующихся к раздражителям и изме |
рвным моделям стимулов. Мотивации захваты |
|
нениям условий среды, которая через снижение |
вают неблагоприятным ФС целые сенсорные |
|
напряжения собственных адаптационных меха |
системы. |
|
низмов способствует адаптации и всей многокле |
ЛИТЕРАТУРА |
|
точной колонии организма. Обеспечение нервной |
||
системой адаптации организма к окружающей |
1. Анохин П.К. Эмоции. Большая медицинс |
|
среде через мобилизацию внутрисистемных меха |
кая энциклопедия. 2 е изд. / П.К. Анохин. –1964. |
|
низмов осуществляется посредством генерации |
– Т. 35. – С. 339. |
|
нейронами ПД. |
|
2. Батуев А.С. Функции двигательного анали |
Рассмотрение ФС нейронов мозга как показа |
затора / А.С. Батуев. – Л.: ЛГУ, 1970. –224 с. |
|
теля эффективности адаптационных механизмов |
3. Беллер Н.Н. Организация и механизмы цен |
|
позволяет по новому взглянуть на физиологичес |
тральных эфферентных влияний на висцеральные |
|
кий механизм многих психических феноменов и |
функции / Н.Н. Беллер. – Л.: Наука, 1983. – 35 с. |
|
их проявление в норме и при патологии, а также |
4. Берталанфи Л.Ф. Общая теория систем / |
|
на природу организма человека и животных в це |
Л.Ф. Берталанфи // Системные исследования. – |
|
лом. Так функциональная роль нервной системы, |
М., 1969. – С. 30–34. |
|
заключающаяся в обеспечении эффективной |
5. Василевская Н.Е. О функции и структуре |
|
адаптации организма, в свете развиваемого пред |
висцерохимического анализатора / Н.Е. Василев |
|
ставления, реализуется через активацию механиз |
ская. – Л.: Изд во ЛГУ, 1971. – 192 с. |
|
мов нейрональной адаптации. Через адаптацию |
6. Васильев Л.Л. Об основных функциональ |
|
нейронов адаптируется и весь организм: хорошо |
ных состояниях нервной ткани / Л.Л. Васильев // |
|
нервным клеткам значит хорошо и всей клеточной |
Новое в рефлексологии и физиологии нервной |
|
колонии, всему организму. |
|
системы. – Л. – М., 1925. – С. 1–41. |
Определение мотиваций, эмоций и внимания |
7. Введенский Н.Е. Возбуждение, торможение |
|
с точки зрения поляризационной теории |
и наркоз / Н.Е. Введенский. – СПб., 1901. –110 с. |
|
Мотивация – это процесс мобилизации ней |
8. Веселовский Н.С. Выделение, идентифика |
|
ронами мозга, находящимися в неблагоприятном |
ция и свойства кальциевого компонента потенци |
|
функциональном (метаболическом) состоянии, |
ала действия в изолированных нейронах спиналь |
|
проявляющемся в устойчивой деполяризации |
ных ганглиев крыс / Н.С. Веселовский, Е.Д. Нали |
|
МП, механизма восстановления их хорошего ФС. |
вайко // Нейрофизиология. – 1986. – Т. 18, № 1. |
|
Эмоция – психический феномен пережива |
– С. 126–129. |
|
ния субъектом изменений функционального (ме |
9. Виноградова О.С. Гиппокамп и память / |
|
таболического) состояния нейронов афферентных |
О.С. Виноградова. – М.: Наука, 1975. – 333 с. |
|
(сенсорных) систем мозга, возникающих при дей |
10. Гельгорн Э. Эмоции и эмоциональные рас |
|
ствии раздражителей из внешней и внутренней |
стройства / Э. Гельгорн, Д. Луфборроу. – М.: |
|
среды организма. |
|
Мир, 1966. – 672 с. |
Ухудшение функционального (метаболическо |
11. Глутамат вызывает понижение мембранно |
|
го) состояния нейронов мозга, выражающееся в их |
го потенциала митохондрий в культивированных |
|
деполяризации, субъективно переживается в виде |
клетках зернах мозжечка / Н.К. Исаев, Д.Б. Зоров, |
|
негативной эмоции. Иначе говоря, негативные |
А.А. Лыжин и др. // Бюл. экспериментальной био |
|
эмоции – субъективное переживание формиро |
логии и медицины. – 1994. – № 2. – С. 208. |
|
вания мотивированного состояния. |
|
12. Голиков Н.В. Работы Л.Л. Васильева об ос |
Улучшение функционального (метаболическо |
новных нервных процессах и их место в совре |
|
го) состояния нейронов мозга проявляющееся в |
менной физиологии / Н.В. Голиков, А.Г. Копылов |
|
реполяризации МП, субъективно переживается в |
// Физиологические механизмы основных не |
|
виде положительной эмоции. |
|
рвных процессов. Труды ленинградского обще |
Превосходное функциональное (метаболичес |
ства естествоиспытателей. –1985. – Т. 75, Вып. 5. |
|
кое) состояние нейронов мозга, отражающее по |
– С. 15–23. |
|
вышение клеточной резистентности и выражаю |
13. Голиков Н.В. Физиологическая лабиль |
|
щееся в гиперполяризации клеток, субъективно |
ность и ее изменения при основных нервных про |
|
переживается в виде сильных положительных |
цессах / Н.В. Голиков. – Л., 1950. – 240 с. |
|
эмоций. |
|
14. Замбржитский И.А. Лимбическая область |
Внимание – разновидность мотивации, |
большого мозга / И.А. Замбржитский. – М.: Ме |
|
представляет собой формирование неблагопри |
дицина, 1972. –280 с. |
|
ятного функционального (метаболического) со |
15. Замбржитский И.А. Пищевой центр мозга |
|
стояния сенсорных нейронов при действии раз |
/ И.А. Замбржитский. – М.: Медицина, 1989. – |
|
дражителей преимущественно из внешней сре |
230 с. |
|
ды. Отличие от мотивации лишь в степени небла |
16. Кулинский В.И. Защитный эффект ин |
|
гоприятного состояния и объеме, захваченной |
терцеребровентрикулярного введения А Агонис |
|
им нервной ткани. Внимание связано с относи |
тов при полной ишемии головного мозга / В.И. Ку |
|
|
|
|
172 |
Экспериментальные исследования в медицине и биологии |
|
|
БЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45) |
|
линский, Л.А. Усов, Г.З. Суфианова, А.А. Суфиа |
29. Мурик С.Э. Об использовании постоянно |
|
нов // Бюл. экспериментальной биологии и меди |
го тока для изучения эмоционального поведения |
|
цины. – 1994. –№ 6. – С. 622–624. |
животных / С.Э. Мурик // В кн.: Актуальные про |
|
17. Леонова А.Б. Психодиагностика функцио |
блемы биологии: Тез. докл. к юбилейной научной |
|
нальных состояний человека / А.Б. Леонова. – М.: |
конф. – Иркутск, 1994. – С. 45. |
|
МГУ, 1984. – 199 с. |
30. Мурик С.Э. Об исследовании постоянно |
|
18. Медведев В.И. Функциональные состоя |
го потенциала головного мозга крыс при пищевом |
|
ния оператора / В.И. Медведев // Эргономика: |
поведении / С.Э. Мурик // Тез. юбилейной научн. |
|
Принципы и рекомендации. – М., 1970. – Вып. 1. |
конф., посвященной 150 тилетию со дня рождения |
|
– С. 127–160. |
И.П. Павлова. – СПб., 1999. |
|
19. Мовчан Н.П. Исследования Л.Л. Васильева |
31. Мурик С.Э. Омегоэлектроэнцефалогра |
|
– новый этап в развитии учения Н.Е. Введенско |
фия – новый метод оценки функционального и |
|
го о парабиозе / Н.П. Мовчан // Физиологические |
метаболического состояния нервной ткани / |
|
механизмы основных нервных процессов. Труды |
С.Э. Мурик // Бюл. ВСНЦ СО РАМН. – Иркутск, |
|
Ленингр. об ва естествоисп. – Л., 1985. – Т. 75, |
2004. – № 1, Т. 3. – С. 189–154. |
|
Вып. 5. – С. 5–15. |
32. Мурик С.Э. Поведение и поляризацион |
|
20. Мурик С.Э. Исследование роли поляриза |
ные процессы в нервной системе / С.Э. Мурик // |
|
ционных процессов головного мозга в механизме |
Тез. ХVII съезда физиологов России. – Ростов на |
|
мотивированного и эмоционального поведения / |
Дону, 1998. – С. 252–253. |
|
С.Э. Мурик // Новое в изучении пластичности |
33. Мурик С.Э. Подход к изучению нервного |
|
мозга: Матер. конф. – М., 2000. – С. 58. |
механизма мотиваций и эмоций / С.Э. Мурик // Ин |
|
21. Мурик С.Э. Мотивации. Эмоции. Внима |
теллектуальные и материальные ресурсы Сибири: |
|
ние: общность нейрофизиологического механиз |
Матер. IV региональной научн. практ. конф. Том «Ес |
|
ма / С.Э. Мурик // Проблемы нейрокибернетики: |
тественные науки». – Иркутск, 2001. – С. 78–82. |
|
Матер. юбилейной международной конф., посвя |
34. Мурик С.Э. Системная организация фун |
|
щенной 90 летию со дня рождения А.Б. Когана. – |
кциональных состояний человека / С.Э. Мурик / |
|
Ростов на Дону, 2002. – С. 186–187. |
/ Интеллектуальные и материальные ресурсы Си |
|
22. Мурик С.Э. Новый подход к изучению не |
бири: Сб. научн. тр. Серия «Естественные науки». |
|
рвного механизма мотиваций и эмоций / С.Э. Му |
– Иркутск, 2002. – С. 160–165. |
|
рик // Матер. XVIII съезда физиологического об |
35. Мурик С.Э. Структурно функциональная |
|
щества им. И.П. Павлова. – Казань, 2001. – |
организация эмоционального мозга (подход к ре |
|
С. 165. |
шению проблемы) / С.Э. Мурик // Организация |
|
23. Мурик С.Э. О влиянии электрической по |
и пластичность коры больших полушарий голов |
|
ляризации зрительной коры на обработку инфор |
ного мозга: Матер. конф. – М., 2001. – С. 56. |
|
мации / С.Э. Мурик // Тез. докл. V выездного за |
36. Мурик С.Э. Экспериментальные данные |
|
седания экспертного совета по молекулярной био |
об электрофизиологических коррелятах ишемии |
|
логии и медицинской биотехнологии посвященно |
мозга разной тяжести / С.Э. Мурик, А.А. Суфиа |
|
го памяти В.М. Бехтерева. – Иркутск, 1990. – |
нов, Г.З. Суфианова, А.Г. Шапкин // Бюл. ВСНЦ |
|
С. 194–195. |
СО РАМН. – 2003. – № 1. – С. 148–154. |
|
24. Мурик С.Э. О поляризационных процес |
37. Насонов Д.Н. Местная реакция протоплаз |
|
сах в анализаторах при эмоциях / С.Э. Мурик // |
мы и распространяющееся возбуждение / Д.Н. На |
|
В кн.: II Съезд физиологов Сибири и Дальнего Во |
сонов. – Л., 1959. |
|
стока. Тез. научных сообщений. – Новосибирск, |
38. Сеченов И.М. Рефлексы головного мозга |
|
1995. – С. 305–306. |
(1866 г.) / И.М. Сеченов // Избранные произведе |
|
25. Мурик С.Э. О роли поляризационных про |
ния. – М., 1953. – С. 31–117. |
|
цессов головного мозга в анализе сенсорной ин |
39. Симонов П.В. Мотивированный мозг / |
|
формации и организации поведения / С.Э. Мурик |
П.В. Симонов. – М.: Наука, 1987. – 269 с. |
|
// III Съезд физиологов Сибири и Дальнего Восто |
40. Симонов П.В. Эмоциональный мозг / |
|
ка: Тез. докл. – Новосибирск, 1997. – С. 155–156. |
П.В. Симонов. – М.: Наука, 1981. – 140 с. |
|
26. Мурик С.Э. О связи поляризационных |
41. Скребицкий В.Г. Регуляция проведения |
|
процессов в анализаторах с эмоциями / С.Э. Му |
возбуждения в зрительном анализаторе / В.Г. Скре |
|
рик // В кн.: Сборник научных работ, посвящен |
бицкий. – М.: Медицина, 1977. –160 с. |
|
ный 75 летию кафедры нормальной физиологии |
42. Сологуб М.И. Функциональные характе |
|
ИГМУ. – Иркутск, 1996. – С. 73–78. |
ристики клеток при их гиперполяризации и депо |
|
27. Мурик С.Э. О связи эмоций с поляризаци |
ляризации / М.И. Сологуб // Физиологические |
|
онными процессами в анализаторах / С.Э. Мурик |
механизмы основных нервных процессов: Тр. Ле |
|
// В кн.: Восстановительная неврология 3. Тез. докл. |
нингр. об ва естествоиспытателей. – Л., 1985. – |
|
международн. симп. – М., 1995. – С. 65–67. |
Т. 75, № 5. – С. 31–40. |
|
28. Мурик С.Э. О функциональном состоянии |
43. Способ определения функционального и |
|
нейронов головного мозга / С.Э. Мурик // Бюл. |
метаболического состояния нервной ткани / |
|
ВСНЦ СО РАМН. – Иркутск, 2003. – № 7. – |
С.Э. Мурик, А.Г. Шапкин // Патент на изобрете |
|
С. 51–53. |
ние. – RU № 2245673 С2. – 2005. |
|
|
|
|
Экспериментальные исследования в медицине и биологии |
173 |
|

БЮЛЛЕТЕНЬ ВСНЦ СО РАМН, 2005, ¹ 7 (45)
44.Шапот В.С. Энергетический обмен голов ного мозга и проблема гипоксических состояний
/В.С. Шапот, К.Г. Громова // Биохимия нервной системы. – Киев, 1954. – С. 139–150.
45.Andersen P. Inhibitory phasing of neuronal discharge / P. Andersen, J. Eccles // Nature. –1962.
– Vol. 196. – P. 645–647.
46.Anoxic terminal negative DC shift in human neocortical slices in vitro / R. Kohling, A. Schmi dinger, S. Hulsmann et al. // Brain Res. – 1996. – Nov. 25. – Vol. 741 (1–2). – P. 174–179.
47.Cannon W.B. Hunger and thirst / W.B. Can non // The foundations of experimental psychology. Edited by C. Murchison. – Worcester. Mass.: Clark University Press, 1929. –P. 434–448.
48.Dell P.C. Some basic mechanisms of the translation of bodily needs into behavior / P.C. Dell /
/In: Symposium on the neurobiological basis of be havior. Eds.: G.E.W. Wolstenholme, C.M. O’Conner.
– Boston: Little, Brown, 1958. – P. 187.
49.Human Physiology / Eds.: R. Schmidt, G. Thews. – Vol. 1. – Springer Verlag, 1983.
50.James W. What is emotion / W. James // Mind. – 1884. – N 4. – P. 188.
51.Kristian T. Calcium in ischemic cell death / T. Kristian, B.K. Siesjo // Stroke. – 1998. – Vol. 29, N 3. – P. 705–718.
52.MacLean P. The limbic brain in relation to the pshychoses / P. MacLean // In: Physiological correlates of emotion. – New York – London: Acad. Press, 1970. – P. 129–146.
53.MacLean P. The limbic system (visceral brain) in relation to central gray and reticulum of the brain stem / P. MacLean // Psychosom. Med. – 1955. – Vol. 17. – P. 355–366.
54.Marczynski T.J. Neurochemical interpre tation of cortical slow potentials as they relate to cognitive processes and a parsimonious model of mammalian brain / T.J. Marczynski // In: Slow poten tial changes in the human brain. Eds.: W.C. McCal
lum, S.H. Curry. – New York: Plenum Press, 1993. –
P.253–275.
55.Mattson M.P. Apoptotic and antiapoptotic mechanisms in stroke / M.P. Mattson, C. Culmsee, Z.F. Yu // Cell Tissue Res. – 2000. – Vol. 300, N 1.
– P. 173–187.
56.Morgan C.T. Physiological theory of drive in Psychology: A Study of a Science (Sensory. perceptual and physiological formulations) / C.T. Morgan // Ed.: S. Koch. – New York: McGraw, 1959. – P. 645.
57.Murik S.E. Approach to the study of neuro physiological mechanism of food motivation / S.E. Murik // Intern. J. Neuroscience. – 2002. – Vol. 112. – P. 1059–1072.
58.Murik S.E. Polarization processes in the nervous system and behavior / S.E. Murik // Intern. J. Neuroscience. – 1998. – Vol. 94. – P. 213–221.
59.Murik S.E. Simultaneous recording of the EEG and direct current (DC) potential makes it possible to asses the functional and metabolic state of the nervous tissue / S.E. Murik, A.G. Shapkin // Intern J. Neuroscience. – 2004. – Vol. 114. – P. 921–934.
60.Murik S.E. The relation of emotions to polarization processes in sensory systems / S.E. Murik // Intern. J. Neuroscience. – 1997. – Vol. 88. – P. 185–197.
61.Olds J. Drives, rewards, and the brain / J. Olds. M.E. Olds // In: New directions in physiology.
– Vol. 2. – New York: Holt, 1965. – P. 327–410.
62.Rolls E.T. The brain and emotion / E.T. Rolls.
– Oxford: Oxford University Press, 1999.
63.Stellar E. The physiology of motivation / E. Stellar // Psychol. Rev. – 1954. – Vol. 61. – P. 5–22.
64.Vincenzi F.F. The pharmacological impli cations of calmodulin / F.F. Vincenzi // Trends Pharm. Sci. – 1981. – Vol. 2. – N 9. – P. 7–9.
65.Winer N. Cybernetics (1948) / N. Winer. – М.: Советское радио, 1968.
174 |
Экспериментальные исследования в медицине и биологии |