

Нейрофизиология / Серотанин
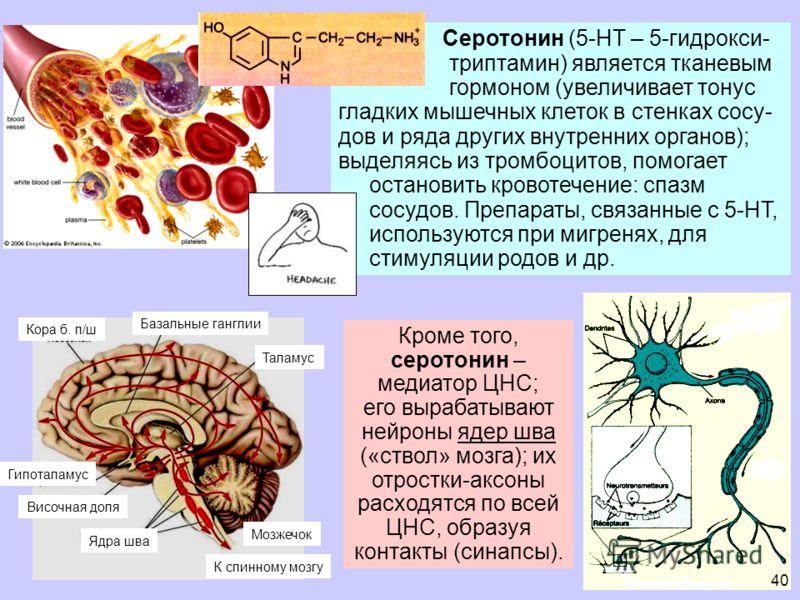
.docxСеротонинергическая система - это нейронная система управления работой мозга, использующая в качестве трансмиттера серотонин. Она образована нейронами, тела которых лежат в ядрах шва продолговатого мозга, тормозит восходящие активирующие системы мозга и тем самым снижает их эффект по активации больших полушарий головного мозга.
Действие третьего поколения антидепрессантов (селективных ингибиторов обратного захвата серотонина - СИОЗС) обусловлено накоплением серотонина в синаптической щели опосредованно через ингибирования систем обратного захвата серотонина. Успех применения данного вида препаратов при различных психических расстройствах свидетельствует о том, что в основе главного звена их патогенеза лежит нарушение функционирования серотонинергической системы, что диктует необходимость подробного рассмотрения функционирования данной эргической системы.
В настоящее время известно около 15 видов серотониновых рецепторов, однако далеко не все их виды и подвиды встречаются в головном мозге человек.
В головном мозге млекопитающих были обнаружены серотониновые рецепторы 5-НТ1. Их выделено 5 подтипов: A, B, D, E, F. Рецепторы представляют собой протеины, содержащие 365 - 422 аминокислотных остатка. Рецепторы посредством ингибиторных Gi-белков сопряжены с аденилатциклазой, поэтому при активации рецепторов активность аденилатциклазы ингибируется.
Подтип 5-НТ1А широко распространен в ЦНС, в особенности в гиппокампе, перегородке, миндалине, то есть в тех областях мозга, которые, как предполагают, участвуют в формировании настроения. Полагают, что именно этот вид рецепторов вовлечен в реализацию антидепрессивного эффекта СИОЗС. В пользу этого предполоежение свидетельствует тот факт, что использование агонистов 5-НТ1А-рецепторов является эффективным методом в лечении депрессий.
При активации соматодендрических 5-НТ1А-ауторецепторов блокируют выход серотонина из пресинаптической мембраны посредством подавления возбудимости нейрона. При использовании в хроническом режиме СИОБС имеет место длительное непрямое стимулирование ауторецепторов, что приводит к их десентизации, что обусловливает повышение уровня экстранейронального серотонина. При одновременном применении антагонистов ауторецепторов (пиндолола) и СИОЗС антидепрессивный эффект значительнее выражен и проявляется быстрее. Известно, что стимуляция постсинаптических рецепторов может вызвать анксиогенный эффект, а стимуляция пресинаптических - анксиолитический.
Подтип 5-НТ1В локализован во фронтальных отделах коры, стриатуме, базальных ганглиях. 5-НТ1В-ауторецепторы у человека не обнаружены. У человек пресинаптические ауторецепторы , являющиеся функциональными аналогами этих же рецепторов крысы и демонстрирующие близкую к ним, но не идентичную, фармакоолгическую чувствительность, получили название 5-НТ1D-рецепторов. Однако в последующем были обнаружены еще два подтипа 5-НТ1D-рецепторов. Согласно новой классификации их обозначили как 5-НТ1В и 5-НТ1D. Изучение поведения крыс-мутантов, у которых полностью отсутствовали 5-НТ1В-рецепторы, показало, что оно характеризуется повышенной агрессивность и склонностью к потреблению алкоголя.
5-НТ1D-рецепторы локализованы в периферических афферентных нервных окончаниях, ганглиях и нейронах ядер тройничного нерва.
5-НТ1С-рецепторы у человека не обнаружены.
В мембранах головного мозга человека идентифицированы 5-НТ1Е- и 5-НТ1F-рецепторы. 5-НТ1F-рецепторы локализованы в периферических нервных окончаниях, ганглиях в системе тройничного нерва.
В головном мозге человека обнаружены 5-НТ2-рецепторы. 5-НТ2А-рецепторы обнаружены в центральной нервнойсистеме, причем их большая плотность обнаружена в пирамидальных нейронах в большинстве регионов коры головного мозга, скорлупе, базальных ганглиях, в меньшем количестве в гиппокампе и хвостатом ядре. Данный вид рецепторов широко распространен в периферических тканях: мочевом пузыре, сосудах, ЖКТ, тромбоцитах, матке. 5-НТ2А-рецепторы у человека опосредуют эффекты галлюциногенов, анксиогенный эффект, участвуют в формировании полового поведения. Предполагают, что данный подтип рецепторов вовлечен в патогенез депрессии и шизофрении (по-видимому эффект галоперидола обусловлен не только блокадой дофаминовых рецепторв, но и серотониновых, так их блокада наступает при концентрации галоперидола 48 мкМ). Посмертные исследования коры головного мозга выявили увеличение числа 5-НТ2А-рецепторов у лиц, страдавших депрессией и покончивших жизнь самоубийством, в то же время количество этих рецепторов в мозге пациентов, страдавших шизофренией, было снижено. В последние года активно обсуждается значение 5-НТ2А-рецепторов для лечения психозов и шизофрении. Сейчас установлено, что многие атипические антипсихотические препараты обладают равной или большей активностью в отношении рецепторов серотонина по сравнению с рецепторами дофамина: так при применение клозапина меньше наблюдается экстрапирамидных расстройств, что обусловлено активацией 5-НТ2А-рецепторов, регулирующих концентрацию дофамина в стриатуме, в результате его количество увеличивается и компенсируется дефицит экстранейронального дофамина.
Существует предположение, что 5-НТ2А-рецепторы могут быть вовлечены в реализацию эффектов антидепрессантов, поскольку появление терапевтического эффекта и снижение числа рецепторов, а также их десентизация нейромедиаторных механизмов, совпадают по времени.
По-видимому, 5-НТ2А-рецепторы вовлечены в регуляцию сна (Физиология сна>>). Антагонисты (ритансерин) улучшает качество сна, увеличивая медленную фазу, а агонисты сокращают быструю фазу сна.
Функция 5-НТ2А-рецепторов может модифицироваться другими рецепторами: агонисты 5-НТ1А-рецепторов угнетают поведенческие реакции, опосредуемые 5-НТ2А-рецепторами.
5-НТ2В-рецепторы участвуют в патогенеза мигрени, так как их стимуляцию вдет к высвобожднеию NО, далее, релаксации сосудов.
5-НТ2С-рецепторы обнаружены в наибольшем количестве в хориоидальном сплетении, гиппокампе, полосатом теле, черной субстанции у крысы. Полагают, что анксиогенные и панические эффекты обусловлены 5-НТ2С-рецепторами. Поэтому при лечение тревожных расстройства и ажитированной депрессии беспокойство субъективно может усиливаться; при дальнейшем применения СИОЗС беспокойство ослабевает, что связано с одной стороны с десентизацией 5-НТ2С-рецепторов, а с другой - с повышением активности 5-НТ1А-рецепторов. Десентизация опосредуется следующим механизмамом: понижающая регуляция - происходит утрата рецепторов плазматической мембраной. Она осуществлятся путем секвестрации рецепторов в клетке путем отделения от аденилатциклазы. Длительное применения клозапина способствует интернализации рецепторов в эндосомах вглубь клетки. Мыши-мутанты, у которых отсутствуют 5-НТ2С-рецепторы, склонны к судорогам, увеличивается прием пищи, нарушаются когнитивные функции.
5-НТ3-рецепторы, 5-НТ4-рецепторы преимущественно располагаются в периферических тканях и ответственны за побочные эффекты СИОЗС.
В последнее время клонированы 5-НТ5-, 5-НТ6-, 5-НТ7-рецепторы, но их физиологические функции еще не известны.
Ниже приведена таблица, которая заключает в себе краткую информацию о рецепторах, их распределении, внутриклеточных механизмах действия, функциях.
|
Подтип |
Локализация |
Эффекторная система |
Функция |
|
5-НТ1А |
Ядро шва |
Ингибирование аденилатциклазы |
Ауторецептор |
|
5-НТ1B |
Черная субстанция |
Ингибирование аденилатциклазы |
Ауторецептор |
|
5-НТ1D |
Сосуды головного мозга |
Ингибирование аденилатциклазы |
Суживание сосудов |
|
5-НТ1E |
Кора, полосатое тело |
Ингибирование аденилатциклазы |
|
|
5-НТ1F |
Головной мозг, периферия |
Ингибирование аденилатциклазы |
|
|
5-НТ2А |
Тромбоциты, гладкие мышцы, кора |
Активация фосфолипазы С |
Агрегация тромбоцитов, сокращение мышц, нейрональное возбуждение |
|
5-НТ2В |
Дно желудка |
Активация фосфолипазы С |
Сокращение |
|
5-НТ2С |
Хориоидное сплетение |
Активация фосфолипазы С |
|
|
5-НТ3 |
Периферические рецепторы |
Ионный механизм (образование каналов – увеличение проницаемости натрия и калия) |
Нейрональное возбуждение, высвобождение сротонина. |
|
5-НТ4 |
Гиппокамп, ЖКТ |
Активация аденилатциклазы |
Нейрональное возбуждение. Высвобождение ацетилхолина. |
Участие серотонина в деятельности центральной нервной системы многообразно. Это прежде всего обусловлено тем, что оно сопровождается изменениями метаболизма в сторону снижения потребления мозгом глюкозы, поглощения кислорода, лактатов и неорганических фосфатов, а также нарушением соотношения натрия и калия. Установлено возбуждающее действие серотонина на парасимпатический отдел ствола головного мозга и лимбической зоны коры. Он активирует бульбарный отдел ретикулярной формации, но тормозит передачу импульсов через зрительный бугор, мозолистое тело и синапсы коры больших полушарий головного мозга. Кроме того, имеются свидетельства влияния серотонинергической системы мозга на возбудимость вазомоторных и терморегулирующих центров, а также рвотного центра. Согласно современным представлениям, серотонин играет основную роль в регуляции настроения. С нарушением функции серотонинергической системы связывают развитие психических нарушений, проявляющихся депрессией и тревогой. Избыток серотонина обычно вызывает панику, недостаток вызывает депрессию. Дефицит моноаминов, к каковым относится серотонин, способен приводить к нарушению синаптической передачи в нейронах лимбической системы и формировать депрессивные состояния, протекающие в виде разнообразных клинически очерченных синдромов. Биохимические исследования позволили понять, почему ряд пищевых продуктов может служить своеобразным лекарством от депрессии. При эмоциогенном пищевом поведении, когда пациенты едят для того, чтобы улучшить настроение, уменьшить тоску и апатию, они предпочитают легкоусвояемую углеводную пищу. Повышение поступления углеводов приводит к гипергликемии и вслед за ней к гиперинсулинемии. В состоянии гиперинсулинемии изменяется проницаемость гематоэнцефалического барьера для аминокислоты триптофана - предшественника серотонина, следовательно, увеличивается синтез последнего в центральной нервной системе. Прием пищи может являться своеобразным модулятором уровня серотонина в центральной нервной системе - повышение его синтеза, связанное с поглощением углеводной пищи, приводит одновременно к усилению чувства насыщения и к уменьшению депрессивных проявлений. Тем самым было наглядно показано: булимия и депрессия имеют общий биохимический патогенетический механизм - дефицит серотонина. Серотонинергическая система имеет отношение к различным видам социального поведения (пищевого, полового, агрессивного) и эмоциям. Нейроэндокринные ритмы, настроение, сон, аппетит и когнитивные функции модулируются серотониновой системой среднего мозга. Серотониновая система другой части мозга – префронтальной коры – нарушается при различных видах асоциального поведения (ауто- и экстероагрессия, убийство). Считается, что истощение серотониновой системы префронтальной коры обуславливает поведенческую расторможенность. Изучение содержания серотонина в крови показало более широкие границы колебания его содержания у больных шизофренией по сравнению с другими больными и с психически здоровыми лицами. Серотонинергическая система и суицид. Во многих исследованиях было также продемонстрировано снижение в ткани мозга самоубийц уровня 5-гидроксииндолуксусной кислоты. Это послужило основанием для гипотезы, согласно которой торможение метаболического оборота серотонина в некоторых отделах мозга, в частности, в стволовых структурах и префронтальной коре, является одним из нейробиологических механизмов формирования суицидального поведения. На сегодняшний день серотониновая система наиболее изучена с этих позиций, и все авторы сходятся в том, что дефицит серотонинергической медиации является важным механизмом суицидального поведения. У жертв суицида и у лиц с высоким риском суицида, вероятнее всего, имеет место локальное снижение серотониновой медиации, сопровождающееся повышением активности соответствующих постсинаптических рецепторов. Одним из важных подтверждений этой точки зрения является эффективность антидепрессантов - блокаторов обратного захвата серотонина при депрессиях с суицидальными попытками. Серотонинергическая система и боль. Существенное значение придается серотонину в деятельности антиноцицептивной системы, центральной регуляции болевой чувствительности. Снижение его содержания приводит к ослаблению анальгетического эффекта, понижению болевых порогов, большей частоте развития болевых синдромов. От содержания серотонина в ЦНС зависит и степень выраженности болеутоляющего действия морфина и других наркотических анальгетиков. Полагают также, что анальгетическое действие серотонина может опосредоваться эндогенными опиатами, поскольку он способствует высвобождению бета-эндорфина из клеток передней доли гипофиза. Местное (например, внутримышечное) введение экзогенного серотонина вызывает сильную боль в месте введения. Предположительно серотонин наряду с гистамином и простагландинами, раздражая рецепторы в тканях, играет роль в возникновении болевой импульсации из места повреждения или воспаления. Серотонинергическая система и половое поведение. Серотонинергическая система мозга участвует в регуляции сексуального поведения. Установлено, что повышение уровня серотонина в мозге сопровождается угнетением половой активности, а снижение его содержания ведет к ее повышению. Влияние серотонина на функции некоторых эндокринных желез обусловлено, по-видимому, не только его прямым действием, но и центральными механизмами, так как в подбугорной области мозга обнаружены терминали серотонинергических нейронов, стимуляция которых сопровождается усилением выделения кортиколиберина и соматотропного гормона. Важным является и то обстоятельство, что серотонин стимулирует секрецию адреналина и норадреналина в мозговой части надпочечников. Вероятнее всего это осуществляется также через гипоталамо-гипофизарную систему. Расстройство цикла сон - бодрствование при депрессии связано также с дисметаболизмом серотонина. Он регулирует дельта-сон, инициирует фазу быстрого сна. Нарушения сна могут быть как основной (иногда единственной) жалобой, маскирующей депрессию, так и одной из многих. Это особенно отчетливо видно на примере так называемой скрытой (ларвированной) депрессии (депрессии без депрессии), поскольку при данной форме патологии расстройства сна могут быть ведущим, а порой и единственным проявлением заболевания. Серотонинергическая система и алкоголизм. При оценке предрасположенности к алкоголизму, особое внимание уделяется анализу генетического полиморфизма серотонинового рецептора подкласса 2А (5-НТ2А), так как серотонин участвует в регуляции потребления алкоголя. Прием алкоголя усиливает высвобождение катехоламинов и изменяет концентрацию опиоидов, приводит к временной активации системы подкрепления, что вызывает положительную эмоциональную реакцию. У человека ген 5-НТ2А находится на длинном плече 13-й хромосомы в локусе q14—q21 и характеризуется рядом полиморфизмов в кодирующей области, из которых диаллельный полиморфизм (1438 G/A) в промоторной области рассматривается в качестве генетического маркера, сцепленного с нервно-психическими заболеваниями, в том числе со злоупотреблением алкоголя. Серотонинергическая система и мигрень. Было выявленно, что колебания уровня серотонина в плазме коррелируют с динамикой приступа мигрени и была сформулирована «серотониновая гипотеза» мигрени. В ее патогенезе и механизмах действий антимигренозных средств принимают участие лишь некоторые специфичные подтипы 5-НТ1-рецепторов, локализованных в церебральных сосудах и сенсорном ядре тройничного нерва. Показано, что нейроны серотонинергического дорсального ядра шва (одна из основных структур эндогенной антиноцицептивной системы) и норадренергического голубого пятна ствола имеют многочисленные проекции к сосудам головного мозга и спинальному ядру тройничного нepвa. Установлено, что на пресинаптических окончаниях тройничного нерва локализованы 5-НТ1D-рецепторы и рецепторы к эндотелину. Они находятся за пределами гематоэнцефалического барьера, а их активация приводит к ингибированию выделения нейропептидов кальцитонина, субстанции Р и к предупреждению развития нейрогенного воспаления. Согласно этой концепции, при мигрени (форма асептического нейрогенного воспаления) триггерный фактор предположительно нейрогенной или гормональной природы антидромно активирует периваскулярные афферентные терминали тройничного нерва. Это вызывает деполяризацию нервных окончаний и выделение из них мощных вазодилатирующих и алгогенных веществ - нейропептидов кальцитонина, субстанции Р, нейрокинина A и вазоинтестициального пептида . Данные нейропептиды вызывают расширение сосудов, увеличение проницаемости сосудистой стенки, пропотевание белков плазмы и форменных элементов крови, отек сосудистой стенки и прилегающих участков твердой мозговой оболочки, дегрануляцию тучных клеток, агрегацию тромбоцитов. Конечным результатом нейрогенного воспаления и является боль. Увеличение содержания свободного серотонина плазмы в фазу приступа мигрени связывают с распадом тромбоцитов. Очаговая неврологическая симптоматика, характерная для этого этапа мигренозного приступа, возникает вследствие сужения церебральных сосудов и снижения кровотока в отдельных участках мозга. В фазу головной боли наблюдается увеличение экскреции серотонина и его метаболитов с мочой и последующее снижение его содержания в плазме и спинномозговой жидкости. Это приводит к снижению тонуса церебральных сосудов, их избыточному растяжению, периваскулярному отеку, раздражению болевых рецепторов. Есть основание полагать, что у больных мигренью имеется генетически обусловленный дефект обмена серотонина, который может быть обусловлен многими факторами, в том числе нарушением метаболизма тромбоцитов, дефицитом фермента, разрушающего тирамин в желудочно-кишечном тракте (это подтверждается наличием заболеваний желудочно-кишечного тракта у значительного числа лиц, страдающих мигренью). В безболевом периоде мигрени выявлено повышение чувствительности серотониновых и норадреналиновых рецепторов сосудистой стенки. Внутри сосуда активируется агрегация тромбоцитов, что сопровождается выделением серотонина. Снижается содержание моноаминоксидазы, что также приводит к асептическому нейрогенному воспалению сосуда. Серотонинергическая система и эпилепсия. Одним из нейрохимических механизмов формирования эпилептической активности является изменение обмена триптофана - «утечка» его окисления в центральной нервной системе с серотонинового на кинурениновый путь. В результате в головном мозге снижается уровень серотонина (тормозного нейромедиатора) и возрастает уровень кинуренина, который повышает возбудимость нейронов мозга. Однако, установлено, что серотонин предупреждает у мышей развитие судорог, вызываемых кислородом. Более того, будучи введенным в сонную артерию, он может прекратить развившиеся судороги. Некоторые противосудорожные препараты (фенобарбитал, дилантин и др.) повышают концентрацию серотонина в мозге. Известно и собственно противосудорожное действие серотонина. Он удлиняет положительность сна, вызванного барбитуратами. Особенно выраженное тормозящее действие серотонин оказывает на кору больших полушарий. Тормозящий эффект серотонина обусловлен его непосредственным влиянием на синапсы мозга. Важно то, что, оказывая тормозящее влияние на кору больших полушарий и вовлекающую систему зрительного бугра, серотонин не подавляет активности ретикулярной формации среднего мозга. Не менее выраженным является его свойство избирательно возбуждать подкорковые структуры, связанные с реакцией пробуждения. Серотонину присуща способность активировать холинэстеразу головного мозга, благодаря чему он является не только химическим медиатором, но и модификатором действия ацетилхолина. Серотонинергическая система и нарушение мозгового кровообращения. Известно, что серотонинергические нейроны шва среднего мозга иннервируют церебральные сосуды и их активность влияет на интенсивность мозгового кровотока. Наиболее отчетливые сдвиги наблюдаются при церебральных инсультах. Экспериментальные данные и клинические исследования свидетельствуют о возможном участии серотонина в патогенезе острых нарушений мозгового кровообращения, в частности ишемических инсультов. В этом плане следует учитывать ангиоспастические эффекты серотонина, реализуемые опосредованно через гипоталамус и при непосредственном воздействии на морфологически измененные сосуды мозга. Этому, по-видимому, предшествует изменение содержания серотонина в веществе мозга. Установленное значительное повышение содержания серотонина в спинномозговой жидкости больных субарахноидальным кровоизлиянием, осложненным «отсроченным» вазоспазмом с развитием инфаркта мозга, cвидетельствует о несомненном участии этого биогенного амина в вазоконстрикторном эффекте в отношении церебральных сосудов. Серотонинергическая система и иммунная система. Имеются данные об участии серотонинергической системы в регуляции иммуногенеза. Изменение уровня серотонина существенно влияет на патогенез ряда аутоиммунных заболеваний нервной системы, в частности рассеянного склероза. В последнее время сформировалось направление исследований, направленных на изучение состояния серотонинергической системы у таких больных, и показано, что она существенно изменена. Дефицит серотонина обнаружен в плазме крови больных рассеянным склерозом, у них существенно нарушено состояние тромбоцитарной серотонинергической системы, страдает активный транспорт серотонина тромбоцитами в связи со снижением скорости его обратного захвата. О нарушении серотонинергической системы при рассеянном склерозе также свидетельствуют стойко сниженное содержание лимфоцитов, несущих специфические рецепторы к серотонину, а также низкий титр противосеротониновых антител. Серотонин участвует в процессах аллергии и воспаления. Он повышает проницаемость сосудов, усиливает хемотаксис и миграцию лейкоцитов в очаг воспаления, увеличивает содержание эозинофилов в крови, усиливает дегрануляцию тучных клеток и высвобождение других медиаторов аллергии и воспаления. Серотонин играет важную роль в процессах свёртывания крови. Тромбоциты крови содержат значительные количества серотонина и обладают способностью захватывать и накапливать серотонин из плазмы крови. Серотонин повышает функциональную активность тромбоцитов и их склонность к агрегации и образованию тромбов. Стимулируя специфические серотониновые рецепторы в печени, серотонин вызывает увеличение синтеза печенью факторов свёртывания крови. Выделение серотонина из повреждённых тканей является одним из механизмов обеспечения свёртывания крови по месту повреждения. Также большое количество серотонина производится в кишечнике. Серотонин играет важную роль в регуляции моторики и секреции в желудочно-кишечном тракте, усиливая его перистальтику и секреторную активность. Кроме того, серотонин играет роль фактора роста для некоторых видов симбиотических микроорганизмов, усиливает бактериальный метаболизм в толстой кишке. Сами бактерии толстой кишки также вносят некоторый вклад в секрецию серотонина кишечником, поскольку многие виды симбиотических бактерий обладают способностью декарбоксилировать триптофан. При дисбактериозе и ряде других заболеваний толстой кишки продукция серотонина кишечником значительно снижается. Массивное высвобождение серотонина из погибающих клеток слизистой желудка и кишечника при воздействии цитотоксических химиопрепаратов является одной из причин возникновения тошноты и рвоты, диареи при химиотерапии злокачественных опухолей. Аналогичное состояние бывает при некоторых злокачественных опухолях, эктопически продуцирующих серотонин. Большое содержание серотонина также отмечается в матке. Серотонин играет роль в паракринной регуляции сократимости матки и маточных труб и в координации родов. Продукция серотонина в миометрии возрастает за несколько часов или дней до родов и ещё больше увеличивается непосредственно в процессе родов. Также серотонин вовлечён в процесс овуляции - содержание серотонина (и ряда других биологически активных веществ) в фолликулярной жидкости увеличивается непосредственно перед разрывом фолликула, что, по-видимому, приводит к увеличению внутрифолликулярного давления. Серотонин оказывает значительное влияние на процессы возбуждения и торможения в системе половых органов.